
Volvox
| Volvox | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Volvox aureus |
||||||||||||
| Systematics | ||||||||||||
|
||||||||||||
| Scientific name | ||||||||||||
| Volvox | ||||||||||||
| L. , 1758 |
Volvox or globular algae (from Latin volvere : wälzen , roll) is a genus of multicellular green algae from the class of Chlorophyceae that live in fresh water . The individual cells of Volvox resemble unicellular green algae, which is why Volvox is considered to be an organism close to the threshold from being unicellular to multicellular. Since the mother organisms die when they release the daughter organisms, Volvox is also one of the simplest organisms where death is part of the normal life cycle. By contrast, single-cell organisms can usually reproduce indefinitely through cell division.
features

Volvox forms spheres 0.15 to 1 millimeter in diameter. The cells form a hollow sphere with a colorless jelly inside. In some species, the cells are connected to one another by cytoplasmic cords. An individual contains up to several thousand individual cells. With Volvox globator there are up to 16,000 cells. Each individual cell has two flagella , an ophthalmic spot and a chloroplast as well as two contractile vacuoles , which makes them resemble the green algae Chlamydomonas .
The locomotion takes place with rotation around the longitudinal axis. Polarity results from cell differentiation: in the front area there are only somatic cells , as such they are not involved in reproduction. The somatic cells are used for the movement of the sphere, for photosynthesis and for the production of the extracellular matrix . In the rear area there are larger generative cells , the so-called gonidia .

Volvox , on the right a burst ball that releases daughter balls. Brightfield image

The same scene as on the left now as a dark-field recording.
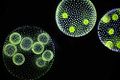
Another darkfield shot.

Another brightfield shot.




Multiplication
Asexual reproduction occurs when the individual gonidia divide synchronously nine to twelve times without increasing their size (growth). The cells are always connected by cytoplasmic bridges, so they form an undivided cell structure ( syncytium ). The resulting embryo already contains all the cells of the future daughter sphere. It forms a single-layer hollow sphere that protrudes inside the nut. The flagella are directed into the interior of the new hollow sphere. In order to create a functional individual, the inside is turned outwards (inversion). Inversion is a complex process of which two variants are known for different types of Volvox . In type A, an opening (called a phialopore) is formed in the hollow sphere. The walls of the sphere turn outward in the shape of a collar and form a bowl-shaped, double-layered stage until they meet again on the opposite side. Then the remaining opening is closed. With type B, the ball bulges inwards from one pole. Only then does a phialopore form on the outside of the indented spherical surface, from which the walls close the sphere again in a rolling movement towards the opposite pole. Changes to the shape, and sometimes also the volume, of the individual cells are essential for the movement processes.
The daughter balls are only released after the mother's somatic cells have died.
Sexual reproduction occurs through oogamy . The gonidia of male organisms form spermatozoids packets through multiple division . These appear as yellowish to orange plates of cells before they are released. The female organisms form uncultivated egg cells . After fertilization , the fertilized egg cell forms an orange permanent zygote with ornamentation on the surface , which can also survive inhospitable times as a persistence stage. The zygote then germinates with reduction division and forms a new individual. However, there are also monoecious ( monoecious ) individuals that form male and female sex cells.
ecology
The genus occurs in fresh water in standing, preferably in eutrophic waters. In particular, all species have a very high phosphate requirement. Volvox species are mostly specialists in very shallow, mostly drying (temporary) shallow waters such as pools and pools. They are also found in layered (stratified) lakes. Volvox species are particularly common in spring and early summer, where they form a separate mass aspect in the annual cycle of eutrophic waters. You can then occasionally form algal blooms . They survive unfavorable or dehydration phases due to the formation of permanent zygospores.
Volvox is often found in heavily turbid waters, which are poorly visible under eutrophic conditions, either due to thrown up mud or due to the high algae biomass. This is where the species benefit from their high mobility, which enables them to always remain in the exposed (euphotic) zone. Volvox is a little heavier than water, so it would sink to the bottom without active movement. The spheres can perceive the direction to the light with their eye spot. In addition, the rear end of the hollow ball is a little heavier, so that the ball tends to swim upwards. Many species can perform diurnal migrations, during which they sink into deeper water layers at night and swim back into the exposed zone during the day. They can cover distances of several meters, which is unusual for phytoplankton and actually more typical for zooplankton. Nutrient uptake (especially phosphate) in deep water is assumed to be the reason for the migration.
Systematics
Volvox is the eponymous genus of the Volvocaceae family. The genus includes around 20 species, three of which occur in Europe.
- Volvox globator with an average of 10,000 individual cells. Individuals are monoecious, with the male sex cells mature first.
- Volvox aureus with 200 to 3200 single cells, dioecious . Pronounced daily vertical migration: during the day near the surface, at night in deeper layers.
- Volvox tertius : the adult cells are not connected by plasma threads.
With the methods of molecular phylogeny, based on the comparison of homologous DNA sequences, it could be clearly demonstrated that the "genus" Volvox is polyphyletic. This means that individual species assigned to the genus are more closely related to representatives of other genera than to each other. Volvox aureus is one of the representatives of the genus Pleodorina , Volvox gigas belongs to the genus Eudorina , Volvox globator and some related species are close to Platydorina . Only the species group around Volvox carteri showed a closer relationship . Volvox in the classic sense is more like a certain type of building plan (level of organization), which was obviously achieved by representatives of different lineages independently of one another. While the type genus Volvox does not represent a natural unit, the monophyly of the order Volvocales, and also its sister group relationship to Chlamydomonas reinhardtii , was confirmed in the analyzes.
The complete genome of Volvox carteri was sequenced in 2010. It turned out that the genome has a comparable size and a comparable number of protein-coding genes as that of the unicellular relative Chlamydomonas reinhardtii, which has already been sequenced . Around 14,500 proteins are predicted for Volvox carteri . Significant differences to the unicellular relatives were found in the genes for the extracellular structural matrix and in the genes that control the sequence of cell divisions.
There are no fossils of Volvox , and given his physique, these are not to be expected. Using the methods of the molecular clock and a comparison with fossilized sister groups (“ ghost lineage ” calibration), a time span of around 234 million years (in the Triassic ) is determined for the evolution of the multicellular nature of Volvox . This is significantly older than earlier estimates, which were only about 50 million years old.
exploration
The first description of Volvox comes from the famous Dutch microscopy pioneer Antoni van Leeuwenhoek , who described it (at the age of about seventy) in 1700. Van Leeuwenhoek already described the characteristic rolling movement of the Volvox balls in water. He also observed the “birth” of daughter balls through a mother's wall.
See also
Web links
swell
- Karl-Heinz Otto Linne von Berg, Michael Melkonian a. a .: The Kosmos algae guide. The most important freshwater algae under the microscope. Kosmos, Stuttgart 2004, ISBN 3-440-09719-6 .
Individual evidence
- ↑ Harold J. Hoops, Ichiro Nishii, David L Kirk: Cytoplasmic Bridges in Volvox and Its Relatives. In: Madame Curie Bioscience Database [Internet]. Austin, Texas: Landes Bioscience; 2000-. [1]
- ↑ Stephanie Höhn & Armin Hallmann (2011): There is more than one way to turn a spherical cellular monolayer inside out: type B embryo inversion in Volvox globator. BMC Biology 2011, 9:89. [2]
- ↑ CS Reynolds Phytoplankton periodicy: its motivation, mechanisms and manipulation. In: Fiftieth annual report for the year ended 31st March 1982. Ambleside, UK, Freshwater Biological Association, pp. 60-75.
- ^ A b David L. Kirk: Volvox: A Search for the Molecular and Genetic Origins of Multicellularity and Cellular Differentiation. Cambridge University Press, 1998.
- ↑ Hans W. Paerl (1988): Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnology and Oceanography 33 (4, part 2): 823-847.
- ^ Ulrich Sommer & Z. Maciej Gliwicz (1986): Long range vertical migration of Volvox in tropical Lake Cahora Bassa (Mozambique). Limnology and Oceanography 31 (3): 650-653.
- ↑ a b c Heinz Streble , Dieter Krauter : Life in the water drop. Microflora and microfauna of freshwater. An identification book. 10th edition. Kosmos, Stuttgart 2006, ISBN 3-440-10807-4 , p. 164.
- ↑ H. Nozaki (2003): Origin and evolution of the genera Pleodorina and Volvox (Volvocales). Biologia (Bratislava) 58: 425-431.
- ↑ David L. Kirk (2005): A twelve-step program for evolving multicellularity and a division of labor. BioEssays 27 (3): 299-310.
- ↑ Simon E. Prochnik, James Umen, Aurora M. Nedelcu, Armin Hallmann, Stephen M. Miller, Ichiro Nishii, Patrick Ferris, Alan Kuo1, Therese Mitros, Lillian K. Fritz-Laylin, Uffe Hellsten, Jarrod Chapman, Oleg Simakov, Stefan A. Rensing, Astrid Terry, Jasmyn Pangilinan, Vladimir Kapitonov, Jerzy Jurka, Asaf Salamov, Harris Shapiro, Jeremy Schmutz, Jane Grimwood, Erika Lindquist, Susan Lucas, Igor V. Grigoriev, Rüdiger Schmitt , David Kirk, Daniel S. Rokhsar (2010): Genomic Analysis of Organizmal Complexity in the Multicellular Green Alga Volvox carteri. Science 329 (5988): 223-226. doi : 10.1126 / science.1188800
- ^ Matthew D. Herron, Jeremiah D. Hackett, Frank O. Aylward, Richard E. Michod (2009): Triassic origin and early radiation of multicellular volvocine algae. Proceedings of the National Academy of Sciences USA Vol. 106 No. 9: 3254-3258. doi : 10.1073 / pnas.0811205106