
Feeler (biology)

In zoology, antennae or antennae are the articulated pair of extremities that appear on the head of most arthropods and are equipped with sensilla ( sense of touch , smell ).

With the exception of the jaw-claw carriers (Chelicerata) antennae are present in all large groups of the arthropods: in the hexipedes with the insects as well as in crustaceans and the millipedes . Of the hexipods , only the leg taster (Protura) have no antennae. Their function here is taken over by the front legs. The tactile organs on the head section of the Chelicerata are called pedipalps .
The head tentacles of the snail are also called antennae.
Antennae homology
According to anatomical relationships, innervation and development (especially gene expression in the creation in the embryo) the antennae of the hexipedes and the millipedes are homologous to the first antennae of the crustaceans; these three orders together form the mandibular animals . These antennae are considered to be the original extremities of the first true segment of the head, which gives rise to the second section of the brain, the deutocerebrum . The head section in front of it with the eyes and the first brain section, the protocerebrum, does not correspond to a real segment and presumably emerges from the original head flap called acron . Their positionally homologous counterparts among the chelicerates are the chelicerae . In the case of the hexipedes, the second antennae originally present in their ancestors were lost early in their evolution. The corresponding head segment, which no longer carries any extremities, is called the intercalar segment.
The antennas of the first head segment are always uniram, i. That is, they are not based on a split leg with two branches. This is sometimes difficult to see in crustaceans because one or more secondary flagella may be present, which can make the antenna appear branched.
The columbus (Onychophora), which is closely related to the Arthropoda, also has a pair of antennae on its head. But these are not homologous to the antennae of the arthropods, they are outgrowths of the original head lobe. This can be recognized, among other things, by the fact that they are innervated by the first section of the brain, the protocerebrum, not, like the antennae of the arthropods, by the second.
Link antennas
.jpg)
Articulated antennas occur in all arthropods with the exception of insects, in the hexipedes, in the springtails (Collembola) and double tails (Diplura). With the exception of the distal phalanx, they contain muscles in all the limbs that insert into the following limb, which means that each limb can be moved individually.
The whip antennae of insects
Flagellant antennas are found in all insects. They sit down together
- Scapus (basal phalanx, first segment), many times enlarged.
- Pedicellus (reversible link) (second segment), often spherical
- Flagellum (or flagellum), with a different number of, very differently shaped, flagellomeric limbs (flagellomeres, antennomeres).
The members of the antenna whip are also often referred to as segments, but in reality they correspond to separate sections or annuli of a single segment.
The antenna is supplied with hemolymph by a blood vessel running through it . A contractile (contractile) vessel, a so-called antenna heart, is often formed at the antenna base, which pumps the hemolymph into the long and thin appendix. It is also traversed by trachea .
The antennae are set off and movable against the head capsule and against each other by joint membranes with a thinned cuticle . The antenna base (the scapus) sits in a cup-like recess in the hard sclerotized head capsule; it is usually a cup joint that can move in all directions ( ball joint ). The end of the scapus anchored in the head capsule is also called the antennae. Muscles that attach to the head (usually the tentorium inside the head capsule) attach to the scapus. A second pair of muscles connects the scapus and pedicellus. The limbs of the antenna whip, however, have no muscles of their own, these can only be moved by moving the base limbs. Further possibilities of movement, for example the spreading of the limbs of a feeler lobe, are generated hydrostatically by the fluid pressure of the hemolymph inside.
The number of whip limbs is highly variable between different insects, it can also be different in very closely related forms. The brush horn sawfly (Argidae) have only one flagellum, many parasitic wasps (Ichneumonidae) have more than fifty, cockroaches like Periplaneta americana 150 or more. The number of antennae elements is fixed within many groups, so most beetles have eleven antennae elements.
In many groups, the antennae in males and females can be designed differently ( sexual dimorphism ). This is often related to the fact that females of many species secrete sexual attractants ( pheromones ), which the males have to perceive over long distances in order to find a successful partner. In adaptation to this, the surface of the male antennae is then greatly enlarged, so that more olfactory cells can be accommodated on it. Examples of this are Diprionids ( bush horn sawfly ) or many groups of butterflies .
Sensory performances
The sensory performance of the insect antennae is based mainly on sensillae, which are microscopic and invisible to the naked eye on the limbs. An exception is the Johnston organ , a special chordotonal organ that works on the basis of scolopidia . Scolopidia are mechanical sensory organs that are located inside the cuticle, i.e. are invisible from the outside and that register a deformation of the exoskeleton . The organ of Johnston registers changes in the position of the antenna whip relative to the antenna base. With Johnston's organ, insects can perceive currents, for example air currents when flying, or gravity (through the tensile forces of the weight of the antenna flagella).

Depending on the type, the antennae have a wide range of sensory hairs, cones (Sensilla basiconica), bottle or ampoule-shaped sensillae with a large terminal opening, plate-shaped sensillae (Sensilla placodea), sensory scales (Sensilla squamiformia), dome-shaped sensillae, and many more other, equipped. The numerous, differently shaped sensillae have different functions. Most of them often act as chemical sensory organs (chemoreceptors), which are primarily used for remote sensing (sense of smell), but occasionally also as contact receptors (sense of taste). Other sensilles serve as mechanoreceptors, moisture receptors or temperature sensors (thermoreceptors), and it is not uncommon for two or even three sensory services to be combined in one type of sensilla. The endowment with sensillae can be repeated on the various members of the flagellum, or certain senses are concentrated on individual members of the flagellum. In many butterflies, for example, the odor sensors are concentrated on the antennae lobes. The number of sensillae depends in part on the size of the limbs and often on the gender. In male cockroaches of the genus Periplaneta there are more than 250,000 sensillae on each antennae, in peacock spiders of the genus Telea around 65,000 in males and only 13,000 in females.
Forms of insect antennae

Sensors can come in very different forms:
- with antenna bristles (aristat), special antenna shape of the flies . The flagellum limbs have largely receded; an often feathery, bristle-like appendage (called arista) is attached to the usually enlarged first flagellum (i.e. the third antenna element).
- lamellar (lamellate), the anterior flagella are enlarged and form a “leaf club”: for example in scarab beetles
- sawn (serrat): the flagellum limbs have a short one-sided extension, for example in click beetles
- fanned out (flabellat): the flagellum limbs have a long, leaf-shaped extension on one side
- like a string of pearls (moniliform): the limbs of the flagella are narrowed at the base and at the tip, the antennae flagella thereby appears constricted; for example in leaf beetles
- bristle-shaped (setiform, English setaceous ) to
filiform (filiform): the antenna whip is long and thin; for example in cockroaches and ground beetles - kneeled (geniculate): the base part of the antenna (scapus) is enlarged or elongated, the other parts are attached to it at an angle; for example in weevils
- feathered (plumos): the flagellum limbs wear long wreaths of hair, for example in the males of mosquitoes
- combed, comb-shaped (pectinate): the flagellum limbs have a moderately long, unilateral appendage, for example in the males of some click beetles
- club-shaped (clavate): the front limbs of the flagella are clearly set apart from the other limbs, enlarged, for example in bark beetles
- awl-shaped (stylate): the antennae whip is very small and thin compared to the two basic members, for example in cicadas
The antennae of the larvae or nymphs of the insects usually have a different shape . Often they are shorter and more simply built.
Antennae of the crustaceans

The head of the crustacean does not have one, but two pairs of extremities, called antennae.
The front antennas are also referred to as "first antennas" or antennas . An antennae is not constructed as a split bone. Usually they (rarely three) consists of two flagella ( flagella ), the segments on several differently constructed (the antenna base, also called peduncle) sit. The flagella are usually designed the same, but can also differ significantly.
The "second antenna" or Antennae are usually the more conspicuous antenna pair, in some species, they are longer than the body. In the basic plan, an antenna is constructed as a split leg, with the endopodite forming the flagellum and the exopodite being present as an elongated or leaf-shaped scale. In decapods , the exopodite is also known as the scaphocerite. The antennas sit on a five-part antenna base.
Conspicuous Antennae have about the lobsters , the antennal Peduncle is considerably increased, or the lobsters in which the scourge of Antenna is transformed into a kind of shovel (see gallery)..
gallery
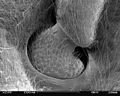
Anchoring of the antenna in the head capsule of the common wasp ( Vespula vulgaris )

kneeling antennae of an ant ( Dorylus )

The feathery antennae of a tufted mosquito (Chaoboridae)

culled antennae of the C-folder

filiform antennae of a cockroach

Greatly enlarged antenna base in lobsters

Reddish antennas and scoop-shaped antennas in slipper lobsters






.jpg)
Individual evidence
- ^ Gerhard Scholtz, Gregory D. Edgecombe: The evolution of arthropod heads: reconciling morphological, developmental and palaeontological evidence . In: Development Genes and Evolution , 216, 2006, pp. 395-415, doi: 10.1007 / s00427-006-0085-4 .
- ↑ Javier Ortega-Hernandez, Ralf Janssen, E. Graham Budd: Origin and evolution of the panarthropod head. A palaeobiological and developmental perspective . In: Arthropod Structure & Development , Volume 46, No. 3, 2016, pp. 354–379, doi: 10.1016 / j.asd.2016.10.011 .
- ^ B. Eriksson, N. Tait, G. Budd, R. Janssen, M. Akam: Head patterning and Hox gene expression in an onychophoran and its implications for the arthropod head problem . In: Development, Genes and Evolution , Volume 220, No. 3-4, 2010, pp. 117-122, doi: 10.1007 / s00427-010-0329-1 .
- ^ Catherine Loudon: Antennae . In: Vincent H. Resh, Ring T. Cardé (Ed.): Encyclopedia of Insects . Elsevier (Academic Press), San Diego 2003, ISBN 0-12-586990-8 .
- ^ Henrik Steinmann, Lajos Zombori: Dictionary of Insect Morphology . Handbuch der Zoologie / Handbook of Zoology Vol. 4, Part 34. Walter de Gruyter, 1999, ISBN 978-3-11-081647-1 .
- ^ Dietrich Schneider: Insect Antennae . In: Annual Review of Entomology 9, 1964, pp. 103-122.
- ↑ Chapter 1: Head . In RF Chapman: The Insects. Structure and Function . Cambridge University Press, 4th edition 1998, ISBN 0-521-57048-4 .
- ^ Gordon Gordh: A Dictionary of Entomology . CABI (Center for Agriculture and Biosciences International) 2011, ISBN 978-1-84593-542-9 .
- ↑ Joel Martin: Antennule. In: Crustacea Glossary. Natural History Museum Los Angeles County, accessed August 17, 2012 .
- ↑ Joel Martin: Scaphocerite. In: Crustacea Glossary. Natural History Museum Los Angeles County, accessed August 17, 2012 .
- ^ Antenna. In: Marine Species Identification Portal - Marine Lobsters of the World - Glossary. Retrieved August 17, 2012 .