
Black sea urchin
| Black sea urchin | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Arbacia lixula on the Mediterranean |
||||||||||||
| Systematics | ||||||||||||
|
||||||||||||
| Scientific name | ||||||||||||
| Arbacia lixula | ||||||||||||
| ( Linnaeus , 1758) |
The black sea urchin ( Arbacia lixula ) is a small benthic sea urchin living in the coastal region ( littoral ). It belongs to the genus Arbacia within the Arbaciidae family.
Naming
Linnaeus first described the black sea urchin in 1758 in its 10th edition of the Systema Naturae as Echinus lixula . The assignment to the genus Arbacia was made in 1835 by John Edward Gray. The generic name is said to be derived from the name Arbaces, a minor character in the historical poem Sardanapalus by Lord Byron, which was published in 1821. According to Harvey (1956), the species name comes from the name of a flat, round biscuit that was made from flour and cheese in ancient Rome and which itself got its name from the Latin verb lixare (Latin: "boil in water").
features
As one of the few (recent) representatives of the Arbacioida order that occurs today, the black sea urchin is initially characterized by features that are common to all "regular" sea urchins ( regularia ). Its radially symmetrical body has a strictly pentameric shape. The mouth and anus of the black sea urchin are located on the two body poles of the round, but slightly flattened body. Furthermore, the hemispherical (hemispherical) inner calcareous skeleton ( testa ) consists of 5 double rows of ambulacral plates. Between these there are 5 double rows of interambulacral plates, which have the spine bases. The paired ambulacral rows are homologous to the arms of the starfish and have pores through which the ambulacral feet open outwards. These plates are rigid, firmly connected to one another and end aborally at the anal area ( periproct ) where the anus opens. The five terminal plates (end plates) of these double rows form a solid ring around the anal field. At the terminal porus of the Testa, the radial channels of the hydrocoel emerge through these terminal plates . On the oral side, the calcareous plates of the testa form a ring around the mouth area (peristome), from the center of which protrude the teeth of Aristotle's lantern , the sea urchins' chewing apparatus. This consists of 5 arrow-shaped, radial pyramids ("jaws"), each of which has a permanently growing tooth. This complex is suspended in the peristome by muscles and ligaments that extend from the inner edge of the peristome and act as lifters and lowerers. Not only do they guide the teeth themselves, but they can also pull the entire lantern out of the mouth and pull it in or move it back and forth within the mouth opening. In the relaxed state, the teeth protrude slightly from the mouth opening. The peristome carries a soft layer of connective tissue from which the black sea urchins' mouth feet emerge. These are ambulacral feet that have small suction cups at their ends. These are used by the black sea urchin to adhere to preferably hard and smooth substrates. The primary spines of this species are evenly arranged densely in rows along the ambulacra and interambulacra. They are brittle and very pointed, about 3 cm long and anchored to the non-perforated tubercles (articular cusps) of the testa with a ligament of connective tissue through which they are moved. They can be erected immediately when touched and serve to protect against predators as well as for locomotion. However, this species has no secondary spines. Since the ambulacral feet on the aboral side of the black sea urchin are not equipped with suction cups compared to other representatives of the regularia, it is very restricted in its locomotion and does not have the ability to mask itself, i.e. H. living animals do not carry any foreign bodies on their laboratory side. The typical stalked, trifoliate grasping arms ( pedicellarians ) are common to all Echinodermata and serve the A. lixula to keep the body clean d. H. for example, to remove small particles or larvae of other marine animals that nestle between the spines. Overall, the bilaterally symmetrical , orally flattened testa has a maximum diameter of 6 cm and, with the exception of the spines, is covered by a black epithelium. The bare testa is reddish to violet-brown, whereby the ambulacras are emphasized by their position between two darker stripes, which also indicate the position of the pores. The living individual is characterized by its dark brown to deep black color. Often it can be observed that the spines on the oral side are more light brown in color. Likewise, smaller individuals are usually characterized by color. The construction of the gonads of the black sea urchin is very typical of all Eleutherozoa . Its hemal and nervous systems are similar to that of starfish.

Aboral aspect of the bare testa of A. lixula

Exterior view of the Lantern of Aristotle

A. lixula in profile

Peristoma of A. lixula




ecology
Habitat and Distribution
The geographical occurrence of A. lixula is limited to Spain and Portugal, the Mediterranean Sea, eastern South America, the Macaronesian Islands, the Atlantic coast and West Africa, with its populations increasing rapidly on the Mediterranean Sea. On the basis of fossil finds , A. lixula is said to have a neotropical origin, since the closure of the isthmus seems to have separated the Pacific species A. stellata from the Atlantic relatives ( A. punctulata / A. lixula ). According to this, A. lixula and the Caribbean species A. punctulata are said to be sister species , which quickly developed away from each other after the closure of this isthmus. Studies also suggest that the black sea urchin, despite its abundance, did not colonize the Mediterranean until the last century. As a representative of the regularia he is a typical inhabitant of the benthos and one of the most common representatives of the sea urchins on coastal rocks of the sublittoral, where he prefers to stay on limestones covered with algae. It can typically be found in shallow water down to a depth of around 30 m, with isolated occurrences at depths of up to 50 m. In addition to the stone sea urchin ( P. lividus), it colonizes bald areas and accumulations of macroalgae, where it prefers to settle on crust-forming calcareous algae and through its diet contributes to the preservation of these bald areas. It is very rarely found in algae forests. So far it is not clear whether the black sea urchin avoids these, as it prefers crust-forming algae as food, or whether the choice of its habitat is based on its inability to move in algae forests. Although this species is not rock-drilling, it can suck itself into the rock in surf zones through the ambulacral feet on its oral side. However, it is not only found on vertical, rocky surfaces where it is exposed to strong hydrodynamics, but also on flat, rocky grounds, broken boulders and rarely even sand.
nutrition
Traditionally, the black sea urchin was regarded as a herbivore based on the analysis of its stomach contents. However, further studies show that this species is an omnivore (omnivore) with a tendency to carnivore . Due to its limited mobility, A. lixula feeds on sessile or very slow, but not on free-swimming ( pelagic ) organisms. When grazing his food he makes use of the teeth of his chewing apparatus to loosen and scrape his food from the substrate. It feeds on various types of brown algae, such as Cystoseira spp. or the representatives of the Dictyotales. However, it prefers to eat crust-forming calcareous algae even in algae forests, which is why it is often said to have selective eating behavior. In addition, A. lixula also feeds on sessile invertebrates when they go pasture, which is why this species is also known as "wolf in sheep's clothing " according to Wangensteen. It can therefore also be assumed that his preference for crust-forming calcareous algae is based on the invertebrates associated with them.
Communities
Many representatives of the species living in the littoral or benthic areas live closely together with A. lixula and seek protection in its immediate vicinity. Young fish of the Gobiidae (gobies), Gobiesocidae (suckers), Blennidae (slime fish ) and Tripterygiidae ( three-fin slime fish ) stay under or in the immediate vicinity of this sea urchin. Crustaceae (crustaceans) such as shrimp, amphipods ( amphipods ) and Paguroidea (hermit crabs) or even polychaetes such as scale worms benefit from the protection between its spines. Mollusks such as beetle snails and front gill snails are mostly found under the black sea urchin.
Predators
Fish are among the most important predators of the sea urchins. So apply Diplodus sargus (SARGO) and Diplodus vulgaris (Diplodus vulgaris) to the most important species among the predators of the Black sea urchin while Coris julis captured only juvenile individuals (sea Young) as representatives of the Labridae (wrasses). Depending on the size ratio between predator and prey, the two Diplodus species make use of different eating habits. Smaller individuals (<1 cm) can be swallowed whole by suction, while larger individuals are detached from the rocky substrate at their base and turned over so that their testa can be broken open by biting into the mouth. Species Chromis chromis (monk fish), Thalassoma pavo (ornate wrasse), Serranus cabrilla (seabass) and Parablennius gattorugine can indeed (Striped blenny) is not the predation be detected per se, but they eat dead specimens of this kind. Especially Chromis chromis can at Eating of the fragments of A. lixula floating in the water can be observed. Since the two representatives of the genus Diplodus, as members of the Sparidae ( sea bream ) family, have an important economic benefit, the overfishing of these species has an impact on the stocks of A. lixula . The effects of these predators and scavengers reach far beyond their direct influence on the populations of A. lixula via so-called trophic cascades and can cause a lasting change in the benthos.
Reproduction and larval development
In A. lixula is a dioecious species whose sexual dimorphism on the differences in length of the genital papillae is limited. The eggs and sperm are abundantly released into the sea water for external fertilization. After the gametes have spawned and subsequently fertilized, A. lixula develops via the Pluteus larva, which is typical of Echinoidea (sea urchins) . It is characterized by its bilaterally symmetrical structure, has 4–6 pairs of arms and its inner skeleton consists of 4 symmetrical and 1–2 unpaired parts. Their planktotrophic phase ends after about 4–6 weeks. It sinks to the bottom and undergoes the metamorphosis to the benthic , juvenile 1 mm sea urchin. This transformation takes a maximum of 1 hour. A. lixula does not care for broods . The success of their settlement is strongly dependent on the suitable substrate. The chemical stimulus from calcareous algae or bare stone make settling easier. According to Fénaux, studies of the gonadosomatic cycle of A. lixula controlled by the photoperiod show that it reaches its peak in the Mediterranean every year from May to June, which correlates with the occurrence of pluteus larvae in plankton between July and November. However, Pedrotti was able to determine the disappearance of the larvae from August to September. Since these are the warmest months and therefore favorable conditions for the development of the larva, a connection between this observation and a low concentration of phytoplankton during these months is assumed. This could increase the mortality rate of the larvae due to the lack of food . Alternatively, a larger occurrence of planktivorous fish is suspected, which the larvae of the black sea urchin free. In artificial insemination experiments, individuals of the black sea urchin can be induced to spawn via a KCL injection. The rearing of the larvae turns out to be problem-free, which is why they serve as model organisms in experimental developmental biology.
Influence of environmental conditions
temperature
A. lixula is considered a thermophilic species. Adult individuals can be found all year round, larvae, depending on the source, especially in August. In addition, a population outbreak can be observed after hot summers. At high temperatures this species shows a higher larval survival rate, which is due to the shortened larval period and the associated reduction in predation. The growth of the larvae is accelerated at temperatures above the average sea temperature of the Mediterranean. This observation suggests that A. lixula resides in suboptimal conditions in the Mediterranean and has actually only recently colonized it. However, this would also mean that a warming of the Mediterranean in the course of global warming would make a previously limiting factor of the A. lixula populations disappear, which could intensify its negative effect on the coastal ecosystem of the Mediterranean in the future.
Salinity
With regard to many species it has already been established that a change in the salinity of their marine habitat affects their survival, reproduction, development or growth. For A. lixula, however, so-called embryotoxicity tests could not demonstrate any influence of a change in salinity on the gametes themselves or on their fertilization. The optimal range of salinity for the development of the black sea urchin is between 29 and 35.5% and thus even covers a higher tolerance range than the related species Paracentrotus lividus.
Nanoparticles
There are numerous studies that deal with the effects of nanoparticles on the environment and organisms. These are used in nanotechnologies in various areas such as medicine, the textile industry or engineering, which means that they often find their way into the environment and can influence the ecosystem. Embryotoxicity tests with CuO nanoparticles (copper oxide) show a concentration-dependent decrease in embryo growth, developmental defects and abnormalities in the skeleton of the pluteus larva of A. lixula. This species shows a decline in contaminated seawater, since its early stages of development are particularly sensitive to contact with CuO nanoparticles. Aggregates of these nanoparticles are taken up by the black sea urchin, its cholinergic and serotonergic system show changes and its neurotransmission is influenced.
Likelihood of confusion
The black sea urchin is often confused with the edible and economically heavily used stone sea urchin ( Paracentrotus lividus ) due to its external similarity and coexistence . Fishermen originally considered them to be male and female individuals of the same species. Their respective Italian names are based on this. The black sea urchin was called riccio maschio and the stone sea urchin was called riccio femina . However, on closer inspection, the living individuals differ significantly from one another. The peristomal membrane is significantly larger and more visible in A. lixula than in P. lividus ; its spine-free mouth field therefore takes up a much larger part of the oral side . In addition, the stone sea urchin has shorter and denser spines and, in contrast to the black sea urchin, also has ambulacral feet on the aboral side of its body with suction cups. It therefore tends to mask what A. lixula is not capable of. The stone sea urchin is rock-drilling, which is why you can often find it in caves on the rock and can also be determined using this characteristic. Furthermore, the lantern of Aristotle from A. lixula is larger, which is seen as its evolutionary adaptation to the durophagy because the herbivore P. lividus feeds on non-crust-forming, softer organisms. Furthermore, the gonadal mass of the black sea urchin is smaller and the stone sea urchin is characterized by a much higher mobility due to the distribution of your ambulacral feet all over the body. For a long time these two species were assigned the same ecological niche and trophic level due to their apparent socialization . More detailed investigations into an interaction have yet to be made, but different substrate and food preferences show that their niche cannot completely overlap and that the omnivorous and partly carnivorous species A. lixula must occupy a higher trophic level. It therefore seems unlikely that these species will compete with one another.

Oral side of the testa of A. lixula (l.) And P. lividus (r.)

The testa of A. lixula (left) and P. lividus (right) in comparison
_und_P._lividus_(r.).jpg)
_und_P._lividus_(r.)_im_Vergleich.jpg)
meaning
Ecologically
.jpg)
A. lixula plays an important role in the littoral ecosystem. Its grazing behavior leads to the destruction of algae forests, as well as the creation and maintenance of bald areas on rocky substrates of shallow marine ecosystems. This species controls the dynamics, structure and structure of populations of macroalgae . The relationship between the density of this species and the expansion of bald areas is clear. A high density of the black sea urchin can make algae disappear completely and thus have a lasting effect on the productivity, structure and function of the ecosystem. Furthermore, it can be observed that the number of individuals of limpets of the genus Patella decreases significantly when the black sea urchin is removed from its natural habitat and increases again when it is present. Bulleri suspects that the number of these limpet representatives depends on the density of macroalgae that are regulated by A. lixula . The number of individuals of the red algae genus Peyssonnelia increases when A. lixula is removed , which suggests that these representatives of the red algae are also dependent on it. Overall, the regulating effect of the distribution and frequency of different species of the benthic community ascribes a significant role to the black sea urchin in the marine ecosystem. The reduction in its predators due to changing environmental conditions and human fishing behavior, as well as the tendency towards water warming, which favor the reproductive success of the thermophilic species, increasingly reinforce its ecological role. But this kind of disadvantageous processes, such as the acidification of the oceans associated with global warming , must also be taken into account. Although these do not show any significant influence on adult individuals, they do impair the growth of the larvae. Nonetheless, the ecological importance of this species can only be fully explained in the context of its interaction with the stone sea urchin and therefore needs further research.
Economically

For a long time the black sea urchin was not used economically. However, its gonads are edible and are considered a delicacy in the Mediterranean region that is drizzled with lemon juice and eaten raw. The decline in the more frequently eaten stone sea urchin is gradually bringing it onto the market, although its taste is described as weaker. The individuals are collected by hand and special sea urchin openers serve as tools. However, it should be noted that the gonads of this species can be poisonous during the reproductive season and should therefore not be eaten during this period.
Bio-indicator
Due to its sensitivity to various environmental factors and its availability, A. lixula is considered to be an important bio-indicator for the so-called biomonitoring of the marine ecosystem and therefore also has great ecological significance. Its simple handling and the possibility of keeping it in aquariums favor the possibilities of environmental research based on studies of this species.
Medical
The intense purple color of the eggs of A. lixula is due to the storage of carotenoids , which this species ingests through its diet. This is astaxanthin , an antioxidant that is supposed to support the prevention and treatment of cancer, chronic inflammatory diseases, diseases of the metabolism, the liver, the skin, the eyes or the intestines, diabetes or neurodegenerative diseases. The eggs of the cultivated individuals contain a 15 times higher concentration of this substance than the specimens collected in the natural habitat. As a future source of a substance with pharmaceutical significance, medicine could use the black sea urchin in culture without exhausting its natural populations.

Injury from Arbacia lixula
An injury caused by this type causes unpleasant and sometimes very painful injuries. The spines in the wound usually break off and, although they dissolve within a few days, can cause severe inflammation if the wound is soiled. Contrary to the assumption of many tourists, the spines of the black sea urchin are not poisonous.
Additional images
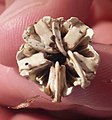
Rear view of the lantern stored in the Testa

Side view of the lantern with its arrow-shaped pyramids

Ambulacral feet of the violet sea urchin (Sphaerechinus granularis) with suction cups

Aboral site of A. lixula


_mit_Saugn%C3%A4pfen.jpg)

Individual evidence
- ↑ a b c d e f g h i Wangensteen, OS (2013). Biology and phylogeography of the black sea urchin Arbacia lixula (Echinoidea: Arbacioida) (Doctoral dissertation, Ph. D. Thesis, Universitat de Barcelona, Spain).
- ↑ a b c d e f g Westheide, W., Rieger, R., Dohle, W., Emschermann, P., Goetting, KJ, Goldschmid, A., ... & Huelsmann, N. (2013). Special zoology. Part 1: Protozoa and invertebrates. Springer Spectrum, Heidelberg.
- ↑ a b c d e f DeHaas, W., Knorr, F. (1999). What lives in the sea on Europe's coasts? Müller Rüschlikon, Stuttgart.
- ↑ a b c Riedl, R. (1983). Fauna and flora of the Mediterranean. P. Parey.
- ↑ a b Tortonese, E., 1965. Echinodermata. Fauna d'Italia vol. VI. Calderini, Bologna.
- ↑ a b c Hickman, CP (2008). Zoology. Pearson Germany GmbH, Hallbergmoos.
- ↑ a b c d Tortonese, E., & Vadon, C. (1987). Oursins et Holothuries (Echinodermes). Fiches d'identification des especes pour les besoins de la peche, 1, 715-739.
- ↑ a b c Carballeira, C., Martín-Díaz, L., & DelValls, TA (2011). Influence of salinity onfertilization and larval development toxicity tests with two species of sea urchin. Marine environmental research, 72 (4), 196-203.
- ↑ Bonaviri, C., Vega Fernández, T., Fanelli, G., Badalamenti, F., Gianguzza, P., 2011. Leading role of sea urchin Arbacia lixula in maintaining barren state in southwestern Mediterranean. Marine Biology 158,2505-2513.
- ↑ a b c d e Agnetta, D., Bonaviri, C., Badalamenti, F., Scianna, C., Vizzini, S., Gianguzza, P., 2013. Functional traits of two co-occurring sea urchins across a barren / forest patch system. J. Sea Res. 76, 170-177.
- ↑ a b Göthel, H. (1997). Lower animals color atlas Mediterranean fauna: lower animals and fish. Ulmer, Stuttgart.
- ↑ Agnetta, D., Badalamenti, F., Ceccherelli, G., Di Trapani, F., Bonaviri, C., & Gianguzza, P. (2015). Role of two co-occurring Mediterranean sea urchins in the formation of barren from Cystoseira canopy. Estuarine, coastal and shelf science, 152, 73-77.
- ↑ Benedetti-Cecchi, L., Bulleri, F., Cinelli, F., 1998. Density dependent foraging of sea urchins in shallow subtidal reefs on the west coast of Italy (western Mediterranean). Mar. Ecol. Prog. Ser. 163, 203-211.
- ↑ a b Hofrichter, R. (2004). The Mediterranean: Fauna, Flora, Ecology - Destination Guide, II / 1. Spectrum Academic Publishing House, Heidelberg.
- ↑ Guidetti, P., Mori M (2005) Morpho-functional defenses of Mediterranean sea urchins, Paracentrotus lividus and Arbacia lixula , against fish predators. Mar Biol 147: 797-802.
- ↑ Guidetti, P., (2004) Consumers of sea urchins ( Paracentrotus lividus and Arbacia lixula ) in shallow Mediterranean rocky reefs. Helgol Mar Res 58: 110-116.
- ↑ a b Privitera, D., Noli, M., Falugi, C., & Chiantore, M. (2011). Benthic assemblages and temperature effects on Paracentrotus lividus and Arbacia lixula larvae and settlement. Journal of Experimental Marine Biology and Ecology, 407 (1), 6-11.
- ↑ Fenaux, L., 1968. Maturation of gonades et saisonnier cycle of Larves chez A. lixula , P. lividus et P. microtuberculatus à Villefranche-sur-Mer. Vie et milieu Series A Biology Marine 19, 1-52.
- ↑ Pedrotti, ML, 1993. Spatial and temporal distribution and recruitment of echinoderm larvae in the Ligurian Sea. Journal of the Marine Biological Association of the United Kingdom 73, 513-530.
- ↑ a b Maisano, M., Cappello, T., Catanese, E., Vitale, V., Natalotto, A., Giannetto, A. & Fasulo, S. (2015). Developmental abnormalities and neurotoxicological effects of CuO NPs on the black sea urchin Arbacia lixula by embryotoxicity assay. Marine environmental research, 111, 121-127.
- ↑ Santos, R., Flammang, P., 2005. Morphometry and mechanical design of tube foot stems in sea urchins: a comparative study. J. Exp. Mar. Biol. Ecol. 315, 211-223.
- ↑ Hereu, B., 2006. Depletion of palatable algae by sea urchins and fishes in a Mediterranean subtidal community. Mar. Ecol. Progr. Ser. 313, 95-103.
- ↑ Sala, E., 2004. The past and present topology and structure of Mediterranean subtidal rocky-shore food webs. Ecosystems 7, 333-340.
- ↑ Bulleri, F., Benedetti-Cecchi, L., & Cinelli, F. (1999). Grazing by the sea urchins Arbacia lixula L. and Paracentrotus lividus Lam. in the Northwest Mediterranean. Journal of Experimental Marine Biology and Ecology, 241 (1), 81-95.
- ↑ Cirino, P., Brunet, C., Ciaravolo, M., Galasso, C., Musco, L., Vega Fernández, T., & Toscano, A. (2017). The sea urchin Arbacia lixula : a novel natural source of astaxanthin. Marine drugs, 15 (6), 187.