
Masrasector
| Masrasector | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
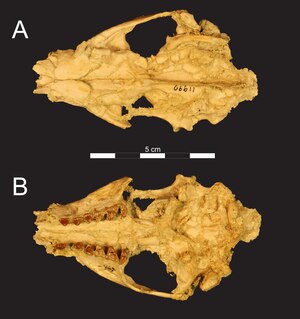
Skull of Masrasector in two views |
||||||||||||
| Temporal occurrence | ||||||||||||
| Upper Eocene ( Bartonian ) to Lower Oligocene ( Rupelian ) | ||||||||||||
| 38 to 30 million years | ||||||||||||
| Locations | ||||||||||||
| Systematics | ||||||||||||
|
||||||||||||
| Scientific name | ||||||||||||
| Masrasector | ||||||||||||
| Simons & Gingerich , 1974 | ||||||||||||
Masrasector is a genus of the order Hyaenodonta , extinct carnivorous mammals that may be close to predators. It occurred in the Upper Eocene and Lower Oligocene 38 to 30 million years ago in what is now Africa and the Arabian Peninsula . The found material consists mainly of skull and dentition remains, and there are also individual elements of the musculoskeletal system. The fossil deposit of Fayyum in Egypt turned out to be the most important find. Small animals can be deduced from the known fossil material. Possibly they again fed on smaller mammals, but specialization in molluscs was also considered. The scientific introduction of the genus dates back to 1974, at present several species have been documented. Initially, the systematic position of Masrasector was controversial due to some special tooth features.
description

Masrasector was a rather small representative of the Hyaenodonta . Small members of the genus had a reconstructed weight averaging 1.05 to 1.16 kg. Larger specimens did not reach the dimensions of the closely related Brychotherium . In terms of size, the animals corresponded to today's Kleinfleck gorse cat or the striped skunk . The skull and individual parts of the musculoskeletal system are of the genus. The known skulls are complete with a length of 8.1 cm, but they show striking deformations caused by positioning. The rostrum was narrow and partly tubular. The middle jaw bone was only a thin bone that formed the lateral edge of the interior of the nose. The nasal bone drew in a little laterally over the nostril. The suture between the nasal and frontal bones formed a V with the point pointing backwards. In the upper jaw, above the third premolar, there was a large infraorbital foramen with a diameter of 3.2 mm. The maxilla itself was not involved in the margin of the orbit . This was formed by the frontal bone at the top, the lacrimal bone in front and the temporal bone at the bottom ; the front edge sat above the first molar . The frontal bone showed a prominent bulge at the upper rear edge of the orbit, but a postorbital process was not developed. The tear bone protruded from the orbit into the face and had a wide area of contact with the upper jaw. The zygomatic arch was quite massive over its entire length. Two prominent temporal lines ran across the frontal bone and served as the insertion of the temporalis muscle . On the parietal bone there was also a strong parietal ridge that reached to the bulge of the occiput . This was like a fan, wide and extensive. The articular surfaces of the occiput for connection to the cervical spine had an oval shape and were thus higher than wide. The skull base is largely damaged. The glenoid groove for the joint of the lower jaw with the skull was also oval with the greatest width across the skull axis.

The lower jaw had a massive structure. The symphysis reached the third premolar and was not closed. The anterior mental foramen was located under the first premolar, two more followed each under the third and fourth premolar. The lower edge of the lower jaw ran convex to about the middle under the crown process. A small concave indentation was formed here, which then ended in a rounded angular process. The mandibular foramen opened below the tip of the crown process . It was oriented horizontally, the mandibular canal attached there measured 2 mm in diameter. The ascending branch of the lower jaw started at an angle of 106 ° to the alveolar plane . It was made broad and loomed high, about 1.25 times the height of the horizontal bone.

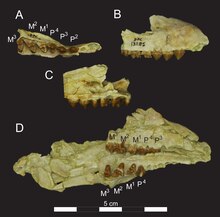
The incisors are not preserved in either the upper or lower half of the dentition . The canine represented the largest tooth, in the lower jaw it reached half the height of the crown process. He was squeezed to the side and curved slightly backwards. There were no sharp cutting edges, so that the cross-section looked more conical than blade- like. All the back teeth were in a closed row. When viewed from the side, the premolars had a solid triangular shape, which was mainly caused by the large paraconus (upper jaw) or the protoconid (lower jaw), the respective main cusps of the premolar teeth. In Masrasector the hump was shaped rather humped and not as pointed as in the related Brychotherium . The rearmost premolars also largely lacked the sharp cutting edges that existed in Brychotherium , for example . The molars had a typical sectorial structure with three main cusps (metaconus, paraconus and protoconus in the upper and metaconid, paraconid and protoconid in the lower molars). The meta- and paraconus of the first two maxillary molars were somewhat laterally pressed, but not as strongly as in Brychotherium . The humps formed a unit at the base, the tips were clearly apart. On the first molar, both cusps were about the same height, on the second the metaconus was slightly higher than the paraconus. The cusps fused at the base and the metaconus, which is higher than the paraconus, are characteristic features of the Teratodontinae, in tribal forms such as dissopsalis and anasinopa the paraconus was also significantly reduced in size. There were sharp cutting ridges on the lateral edges, the posterior (postmetacrista) being connected to the metastyle via a shallow bay; the bay was more prominent on the second molar than on the first. The protoconus was about half as high as the meta- and paraconus. The third maxillary molar was significantly smaller than the previous two and was characterized by the absence of the metaconus. In the lower jaw, the molars had a characteristic talonid (a lower part of the chewing surface) that took up almost half the length of the tooth. As a result, Masrasector differed from Brychotherium with its shorter Talonids and was more like Glibzegdouia . The protoconid formed the largest hump, the smallest the metaconid, while the paraconid mediated between the two. From the anterior to the posterior mandibular molar, the paraconid grew proportionally in size, while the metaconid became smaller. In this characteristic Masrasector agrees with Brychotherium . The largest tooth in the lower jaw was the rearmost molar, 6.2 mm long. In some cases, however, the premolars exceeded the length of the anterior molars. The second premolar was 5.4 mm long, but the first molar was only 4.5 and the second 5.3 mm. The situation was similar in the upper jaw, where the corresponding values for the second premolar were 5.2 mm (the first is unknown), for the first molar 4.7 and for the second molar 5.9 mm. The much smaller third molar was only 2.7 mm long.
Only three fragments of the lower humerus are known. The longest piece is 63 mm long and still has substantial parts of the diaphysis . The shaft was round in cross-section and had a prominent bone ridge as the attachment point for the supinator muscle . The bar narrowed towards the lower joint end ( elbow joint ) and then ran out on the outside on the outer protrusion (epicondyle) of the joint roller. The inner protrusion of the joint roller was enlarged. The olecranon fossa formed a massive opening in the bone above the joint.
Fossil finds
Masrasector is known from several sites in North Africa and the Arabian Peninsula . The most extensive fossil remains include those of the Fayyum region in northern Egypt , one of the most important fossil sites from the Paleogene . The previously known material is stratigraphically scattered relatively widely. The possibly oldest finds here come from the BQ-2 find location in the lower section of the Qasr-el-Sagha Formation and thus belong to the Upper Eocene around 37 million years ago. These are individual isolated teeth and a branch of the lower jaw. The fossil finds from locality L-41 , which is located in the lowest section of the Gebel-Qatrani Formation , are very extensive . The age is about 34 million years. The very fine-grained deposits here differ from the remaining coarser clastic sediments of the rock unit and probably go back to a former lake. Several complete skulls and numerous lower jaws as well as individual long bones have been discovered here. This makes Masrasector one of the smallest hyaenodonts in the Afro-Arab region, from which parts of the musculoskeletal system have been handed down. A few individual finds have been documented from younger layers of the Gebel-Qatrani Formation, which largely date to the Lower Oligocene . A lower jaw remnant, an upper jaw fragment and isolated teeth from site G are to be emphasized . They represent the found material first discovered in research history and were used to establish the genus. Other documented finds of Masrasector received at the sites A , E , I and M . Those of the last two are the most recent finds of the genus in Fayyum.
Apart from the Fayyum region, Masrasector was found in North Africa in Bir el-Ater (Nementcha) in northeastern Algeria . The site with its rich fossil remains belongs to the Upper Eocene, more precisely to the transition from Bartonian to Priabonian . As a result, the finds from Masrasector are probably the oldest of the genus so far. However, they are limited to two upper and one lower molar. Two other teeth came from the Ashawq formation near Taqah in southern Oman . They are of a Lower Oligocene age and should therefore be at the same time as the remains from the upper area of the Gebel Qatrani Formation in the Fayyum.
Paleobiology
Based on the lower joint end of the humerus, a stable elbow joint can be assumed. It can be concluded from this that Masrasector probably moved terrestrially, possibly he was a fast runner. In general, however, hyenodonts have a more massive musculoskeletal system than today's predators. Starting from the low body weight of the animals can be their prey spectrum in Fayyum on the smaller porcupine-like rodents and small representatives of the hyrax constrict. The more tree-dwelling primates are likely to be excluded as a potential food resource. On the other hand, the rather blunt cusps of the premolars could also indicate a preference for hard-shelled molluscs ( durophag ).
Systematics
|
Internal systematics of the Teratodontinae according to Solé & Mennecart 2019
|
Masrasector is a genus from the extinct subfamily of the Teratodontinae within the also extinct order of the Hyaenodonta . The Hyaenodonta were originally considered part of the Creodonta , some of which are sometimes misleadingly referred to as "primal predators". It was assumed that the Creodonta represent the sister group of today's predators (Carnivora) within the parent group of the Ferae . Since the Creodonta subsequently turned out to be a non-self-contained group, they were split into the Hyaenodonta and the Oxyaenodonta . Both groups are distinguished from predators by a further backward shifted in the dentition crushing scissors out. As a rule, the second upper and third lower molars are included in the hyaenodonts. The hyaenodonts can be found for the first time in the Middle Paleocene around 60 million years ago, and they last appeared in the Middle Miocene around 9 to 10 million years ago. The Teratodontinae take within the Hyaenodonta the position of the sister group of the family of Hyainailouridae one, both together form the parent group of Hyainailouroidea . A characteristic feature of the Teratodontinae can be found in the structure of the maxillary molars, in which the para and metaconus are only fused at the base and the latter towers over the former. In contrast, in the Hyainailouridae the Para- and Metaconus merge to form the Amphiconus. In addition, the paraconus is higher than the metaconus. In terms of the height of the two cusps, the Teratodontinae agree with the Hyaenodontidae , but in these the Para- and Metaconus are also fused together. Masrasector is closely related to some of the younger members of the Teratodontinae, such as Dissopsalis , Teratodon and Anasinopa . As a sister genus of the group, Brychotherium can be understood, which is also detected in the Fayyum.
The following types of Masrasector are recognized:
- M. aegypticum Simons & Philip D. Gingerich , 1974
- M. ligabuei Crochet , Thomas , Roger & Al-Sulaimani , 1990
- M. nananubis Borths & Seiffert , 2017
|
Internal classification of the genus Masrasector according to Borths and Stevens 2017
|
Of these three species, M. nananubis is the oldest and smallest representative in phylogenetic terms ; it appeared as early as the Upper Eocene at the L-41 site in Fayyum. The other two species date largely to the Lower Oligocene . M. aegypticum comes from site G of the Fayyum. M. ligabuei was first discovered in Taqah in Oman, but the finds from Bir el-Ater could possibly also represent this species, with which the representative also appeared in the Upper Eocene. Another potential species, " M. pithecodacos ", had already been established in 1994 in an unpublished work by Patricia A. Holroyd for sites I and M of the Fayyum area, but a formally valid description is not yet available. A smaller lower jaw from site G in the Fayyum was initially interpreted as a possible new species, but later interpreted as a young of M. aegypticum ; According to other opinions, it represents a genre not previously described.
Masrasector was first described scientifically in 1974 by Elwyn L. Simons and Philip D. Gingerich . The type material comes from site G of the Fayyum region in Egypt. The holotype (copy number CGM 30978) consists of a left lower jaw with the last premolar and the three molars. The generic name Masrasector is made up of the Arabic word Masr for "Egypt" and the Latin word sector for " Zerschneider " (from secare "to cut up"). In their first description, the authors saw Masrasector in the vicinity of original hyaenodonts such as Proviverra , Sinopa and Metasinopa with an intermediate position between the latter two. They justified this with the, in their opinion, progressive enlargement of the three main cusps of the mandibular molars and the increasing importance of the cutting edges. In the following time, Masrasector, together with other forms such as Anasinopa and Dissopsalis, was therefore repeatedly viewed as "African Proviverrinen", the exact family relationships remained largely unclear. Depending on the weighting of the tooth features, scientists suspected on the one hand a closer connection to similar forms from Southeast Asia ( Kyawdawia and Paratritemnodon ), on the other hand also a close relationship with some American forms ( Sinopa and Prototomus ). The rather blunt and large shape of the premolars, in turn, led some researchers to speculate about a closer relationship with the Teratodon known from the Miocene of Africa , which has so far only been sparsely documented. Phylogenetic studies later confirmed this view and thus led to a reference to the Teratodontinae.
literature
- Matthew R. Borths and Erik R. Seiffert: Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts. PLoS ONE 12 (4), 2017, p. E0173527, doi: 10.1371 / journal.pone.0173527
- Elwyn L. Simons and Philip D. Gingerich: New carnivorous mammals from the Oligocene of Egypt. Annals of the Geological Survey of Egypt 4, 1974, pp. 157-166
Individual evidence
- ↑ a b c d e f g h i Matthew R. Borths and Erik R. Seiffert: Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts. PLoS ONE 12 (4), 2017, p. E0173527, doi: 10.1371 / journal.pone.0173527
- ↑ Matthew R. Borths, Erik R. Seiffert, Katherine Good Berger and Elwyn L. Simons: The oldest Fayum Creodont: Dental and humeral morphology of a new proviverrine hyaenodontid from the earliest Late Eocene of Egypt. Journal of Vertebrate Paleontology 30 (suppl), 2010, p. 63A
- ↑ a b c d e Elwyn L. Simons and Philip D. Gingerich: New carnivorous mammals from the Oligocene of Egypt. Annals of the Geological Survey of Egypt 4, 1974, pp. 157-166
- ↑ a b c d Floréal Solé, El Mabrouk Essid, Wissem Marzougui, Rim Temani, Hayet Khayati Ammar, Mhammed Mahboubi, Laurent Marivaux, Monique Vianey-Liaud and Rodolphe Tabuce: New fossils of Hyaenodonta (Mammalia) from the Eocene (localities of Chambi) Tunisia) and Bir el Ater (Algeria), and the evolution of the earliest African hyaenodonts. Palaeontologia Electronica 19 (3), 2016, p. 41A ( [1] )
- ↑ a b c Jean-Yves Crochet, Herbert Thomas Jack Roger, Sevket Sen and Zaher Al-Sulaimani: Première découverte d'unc créodonte dans la péninsule Arabique: Masrasector ligabuei nov. sp. (Oligocène inférieur deTaqah, Formation d'Ashawq, Sultanate d'Oman). Comptes rendus de l'Académie des sciences. Série 2 311, 1990, pp. 1455-1460 ( [2] )
- ^ A b c Margaret E. Lewis and Michael Morlo: Creodonta. In: Lars Werdelin and William Joseph Sanders (eds.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, pp. 543-560
- ↑ Floréal Solé and Bastien Mennecart: A large hyaenodont from the Lutetian of Switzerland expands the body mass range of the European mammalian predators during the Eocene. Acta Palaeontologica Polonica 64, 2019, doi: 10.4202 / app.00581.2018
- ↑ Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, pp. 1–431 (pp. 122–126)
- ↑ Michael Morlo, Gregg Gunnell, and P. David Polly: What, if not nothing, is a creodont? Phylogeny and classification of Hyaenodontida and other former creodonts. Journal of Vertebrate Paleontology 29 (3 suppl), 2009, p. 152A
- ↑ Floréal Solé: New proviverrine genus from the Early Eocene of Europe and the first phylogeny of Late Paleocene-Middle Eocene hyaenodontidans (Mammalia). Journal of Systematic Paleontology 11, 2013, pp. 375-398
- ↑ Floréal Solé, Eli Amson, Matthew Borths, Dominique Vidalenc, Michael Morlo and Katharina Bastl: A New Large Hyainailourine from the Bartonian of Europe and Its Bearings on the Evolution and Ecology of Massive Hyaenodonts (Mammalia). PLoS ONE 10 (9), 2015, p. E0135698, doi: 10.1371 / journal.pone.0135698
- ↑ a b Matthew R. Borths, Patricia A. Holroyd and Erik R. Seiffert: Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ 4, 2016, p. E2639 doi: 10.7717 / peerj.2639
- ↑ Matthew R. Borths and Nancy J. Stevens: The first hyaenodont from the late Oligocene Nsungwe Formation of Tanzania: Paleoecological insights into the Paleogene-Neogene carnivore transition. PLoS ONE 12 (10), 2017, p. E0185301, doi: 10.1371 / journal.pone.0185301
- ↑ Naoko Egi, Patricia A. Holroyd, Takeshi Tsubamoto, Nobuo Shigehara, Masanaru Takai, Soe Thura Tun, Aye Ko Aung and Aung Naing Soe: A new genus and species of hyaenodontid creodont from the Pondaung Formation (Eocene, Myanmar). Journal of Vertebrate Paleontology 24 (2), 2004, pp. 502-506
- ↑ Naoko Egi, Patricia A. Holroyd, Takehisa Tsubamoto, Aung Naing Soe, Masanaru Takai and Russell L. Ciochon: Proviverrine hyaenodontids (Creodonta: Mammalia) from the Eocene of Myanmar and a phylogenetic analysis of the proviverrines from the para-Tethys area. Journal of Systematic Palaeontology 3 (4), 2005, pp. 337-358
- ↑ Stéphane Peigné, Michael Morlo, Yaowalak Chaimanee, Stéphane Durocq, Soe Thura Tun and Jean-Jacques Jaeger: New discoveries of hyaenodontids (Creodonta, Mammalia) from the Pondaung Formation, middle Eocene, Myanmar - paleobiogeographic implications. Geodiversitas 29 (3), 2004, pp. 441-458
- ↑ Floréal Solé, Julie Lhuillier, Mohammed Adaci, Mustapha Bensalah, Mohammed Mahboubi and Rodolphe Tabuce: The hyaenodontidans from the Gour Lazib area (? Early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae. Journal of Vertebrate Paleontology 12 (3), 2014, pp. 303-322
- ^ Rajendra S. Rana, Kishor Kumar, Shawn P. Zack, Floréal Solé, Kenneth D. Rose, Pieter Missiaen, Lachham Singh, Ashok Sahni and Thierry Smith: Craniodental and Postcranial Morphology of Indohyaenodon raoifrom the Early Eocene of India, and Implications for Ecology, Phylogeny, and Biogeography of Hyaenodontid Mammals. Journal of Vertebrate Paleontology 35 (5), 2015, p. E965308, doi: 10.1080 / 02724634.2015.965308