
Protein kinases
| Protein kinases | ||
|---|---|---|

|
||
| cAMP-dependent protein kinase hexamer, Sus scrofa according to PDB 1CMK | ||
| Enzyme classification | ||
| EC, category | 2.7.-.- , kinase | |
| Response type | Phosphorylation | |
| Substrate | Proteins | |
| Products | Phospho proteins | |
Protein kinases are enzymes that catalyze the transfer of a phosphate group from a donor (usually ATP ) to the side chain hydroxyl (OH) group of an amino acid . Kinases are therefore phospho transferases .
Occurrence of the kinases
According to Gerard Manning, around 2% of all genes in the eukaryotic genome code for kinases. The approximately 518 kinases can 30% of all proteins in a cell phosphorylate . The sum of all kinases in a cell is also called kinom (based on -omik ). A classification based on sequence similarities of the kinase domains, known biological functions, shows 10 groups, 143 families and 212 subfamilies in the kinomes of humans, flies, worms and yeast.
Functions of the kinases
The protein phosphorylation is a very important post-translational control mechanism in the signal transduction of the cell. This includes, for example, the regulation of the activity of enzymes or transcription factors . Phosphorylation can activate / inhibit the target protein or determine its subcellular localization through changes in charge or conformation. The physiological effects of these protein phosphorylations depend on the particular substrate of the kinase.
Protein kinase dysfunction is the cause of numerous diseases. Protein kinases are therefore attractive molecular targets for drug intervention and are being intensively researched by the pharmaceutical industry. “Specific” inhibitors of protein kinases are successfully used in cancer therapy (see, for example, imatinib in the treatment of chronic myeloid leukemia ).
Regulation of kinase activity
Due to their far-reaching influences, the protein kinases themselves must be strictly regulated. Regulatory factors include:
- Cofactors / secondary messenger substances ( Ca 2+ , IP 3 , cAMP etc.)
- Activator and inhibitor proteins
- Pseudo substrates
- Phosphorylation in the active site by
- other protein kinases ( trans- phosphorylation)
- itself ( cis -phosphorylation / auto-phosphorylation)
- Subcellular localization within the cell
Types of protein kinases
Most protein kinases phosphorylate either the amino acids serine / threonine or aromatic tyrosines . However, there are also bispecific protein kinases ( EC 2.7.12 ; e.g. MEK or MAP2K ) that can phosphorylate both serine / threonine and tyrosine residues.
Serine / threonine kinases
These protein kinases ( EC 2.7.11 ) phosphorylate the hydroxyl groups (OH groups) of the amino acids serine and threonine . These kinases are regulated by:
- cAMP or cGMP
- 1,2-diacylglycerine (DAG)
- Ca 2+ or calmodulin
- PIP3 and other phospholipid derivatives
The fact that serine / threonine kinases do not phosphorylate all serines and threonines of other proteins is determined by the interaction with the surrounding peptide sequence. These sequences are called consensus sequences . Due to the low specificity, these kinases do not phosphorylate individual proteins, but entire protein families. These enzymes are inhibited by the fact that a pseudo-substrate binds to the active site by mimicking the target sequence of the corresponding kinase, but has no serine or threonine.
Muscle phosphorylase kinase A
This enzyme ( phosphorylase kinase , EC 2.7.11.19 ) is the first Ser / Thr kinase that was discovered in 1956. It is a key enzyme in glycogen metabolism .
Protein kinase A
This enzyme has several functions in the cell. These include the regulation of glycogen, sugar and lipid metabolism . The protein kinase A ( EC 2.7.11.11 ) is composed of two domains. The smaller one consists of a large number of β-sheets , whereas the large subunit has a large number of α-helices . The catalytic center lies between the two subunits. If ATP binds to a substrate, the subunits twist against each other so that the γ-phosphate group of the ATP comes close to the amino acid to be phosphorylated and the transfer reaction can take place.
You yourself are regulated by cAMP. By binding of cAMP to an inactive tetramer composed of two regulatory and two catalytic subunits (R 2 C 2 ), the regulatory subunits are separated from the catalytic ones, which enables the phosphorylation of other proteins. Protein kinase A is in turn regulated by phosphorylation.
In addition, by activating a phosphodiesterase, the available amount of cAMP is reduced by converting it into AMP. This means that the protein kinase A causes its own inhibition and thus no “permanent activation” of the kinase can occur.
Protein kinase B
The kinase, often referred to as Akt in the English-language literature in particular , is an enzyme of the PI3K / Akt signaling pathway, which has numerous effects on the homeostasis of the cell and regulates survival, apoptosis , proliferation and metabolism . Akt occurs in three closely related isoforms (Akt1, Akt2, Akt3), the sequences of which are encoded on chromosomes 14q32, 19q13 and 1q43, respectively. No differences in substrate specificity could be found between the individual isoforms.
Activation takes place in several steps. Initially, phosphatidylinositol-3-kinases ( PI3K ) are recruited to the membrane by activated receptor tyrosine kinases and thus activated. Activated PI3K catalyze the phosphorylation of phosphatidylinositol to phosphatidylinositol-3-phosphate or of phosphatidylinositol-4-phosphate to phosphatidylinositol-3,4-bisphosphate and of phosphatidylinositol-4,5-bisphosphate to phosphatidylinositol-3,4,5-trisphosphate. The resulting substrates now recruit proteins with Pleckstrin Homology (PH) domains to the membrane. The Akt docked in this way can now be activated via a further signal molecule which has PH domains and which phosphorylates phosphoinositide-dependent kinase 1 (PDK1).
As mentioned above, protein kinase B regulates, among other things, survival, proliferation and the death cycle of the affected cell. One example is the increased occurrence of the glucose receptor GLUT-4 with an incoming insulin signal. The transcription of this carrier is fired up in a cascade-like manner and its incorporation is triggered by vesicle transport and constriction. The activity of the enzyme can itself be regulated, for example, by the tumor suppressor protein PTEN , which is used for the dephosphorylation of, for example, phosphatidylinositol-3,4,5-trisphosphate to phosphatidylinositol-4,5-bisphosphate (or other 3'-phosphorylated phosphoinositides ) responsible is. This makes the substrate of the protein kinase unsuitable because it is no longer able to dock onto the PH domain.
The function and regulation mechanisms are manifold and are still being examined in detail today, since it is assumed that a mutation in certain areas of the signal path (e.g. PTEN) is a basis for the formation of tumors .
Protein kinase C
The term protein kinase C ( EC 2.7.11.13 ) applies to a family of proteins with 12 members in mammals, the Ca 2+ , diacylglycerol , and a phospholipid such. B. need phosphatidylcholine for activation. In most cases, protein kinase Cα is what is meant when these enzymes are mentioned.
The protein kinases are highly conserved proteins that consist of an N-terminal regulatory domain and a C-terminal catalytic domain. As long as the enzyme is not activated by a tumor promoter such as tetradecanoyl phorbol acetate (TPA) or one of the cofactors mentioned above, it is inactive. The general linear structure:
N - Pseudo -substrate - TPA / DAG binding - Ca 2+ binding - ATP binding - Substrate binding - C
During activation, protein kinase C shifts to the cell membrane with the help of receptors for activated protein kinase C ( RACK proteins ). After activation, these kinases remain active for a long time, although the Ca 2+ concentration has decreased again. This is attributed to the action of diacylglycerol , which is formed from phosphatidylinositol by a phospholipase . This is activated by the same signals as the protein kinase itself.
The target sequence of protein kinase C is similar to that of protein kinase A in that it contains many basic amino acid residues near the phosphorylated Ser / Thr residues. Substrates of protein kinase C are MARCKS proteins, MAP kinases , transcription factor inhibitor IκB, vitamin D 3 receptor , RAF kinase , calpain and the EGF receptor .
Tyrosine kinases
The tyrosine kinases ( EC 2.7.10 ) include so-called receptor tyrosine kinases (RTKs) and tyrosine kinases without a receptor function. Around 50 different RTKs are grouped into 18 receptor families. According to the ligands , these RTKs are grouped into the following families:
Multifunctional Kinases
Some kinases phosphorylate metabolites as well as other proteins. Thus, the phosphoglycerate kinase 1 (PGK1) phosphorylates both 3- phosphoglycerate (by means of ATP) to 1,3-diphosphoglycerate and at least 3 proteins, e.g. B. Beclin 1 (involved in autophagy ), PDHK1 (pyruvate metabolism), and Bcl2 ( apoptosis ).
Individual evidence
- ↑ Manning G. Genomic overview of protein kinases. WormBook. 2005 Dec 13;: 1-19. Review. PMID 18050405
- ↑ Krebs EG, Fischer EH. The phosphorylase b to a converting enzyme of rabbit skeletal muscle. Biochim Biophys Acta . 1956 Apr; 20 (1): 150-7. PMID 13315361
- ^ Aaronson SA. Growth factors and cancer. Science. 1991 Nov 22; 254 (5035): 1146-53. Review. PMID 1659742
- ↑ Zhimin Lu, Tony Hunter: Metabolic Kinases Moonlighting as Protein Kinases . In: Trends in Biochemical Sciences . tape 43 , no. 4 , April 2018, ISSN 0968-0004 , p. 301-310 , doi : 10.1016 / j.tibs.2018.01.006 , PMID 29463470 , PMC 5879014 (free full text) - ( elsevier.com [accessed on July 22, 2018]).
Web links
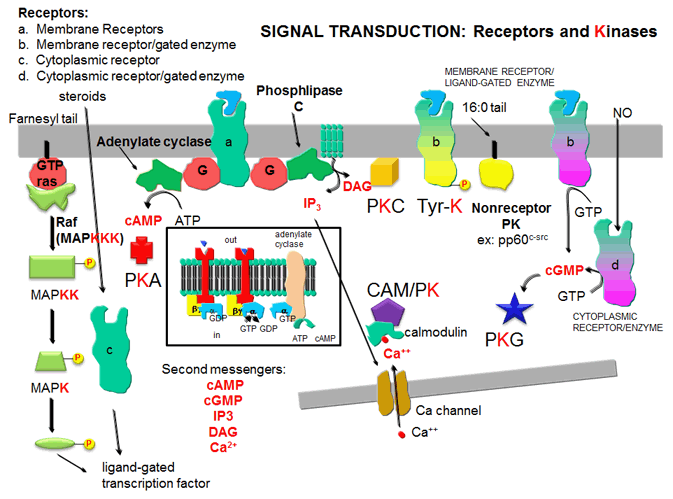
- Receptors and Kinases: five major protein kinases - representation of the role of major protein kinases in signal transduction (English)
- The Protein Kinase Resource - Comprehensive presentation of the kinome with interactive options for comparing individual kinases with regard to function and structure
{kind=link}