
Pampatheriidae
| Pampatheriidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Reconstruction of the skeleton with tanks from Holmesina |
||||||||||||
| Temporal occurrence | ||||||||||||
| Middle Miocene to Lower Holocene | ||||||||||||
| 18 million years to 10,000 years | ||||||||||||
| Locations | ||||||||||||
| Systematics | ||||||||||||
|
||||||||||||
| Scientific name | ||||||||||||
| Pampatheriidae | ||||||||||||
| Paula Couto , 1954 | ||||||||||||
The Pampatheriidae are an extinct family from the order of the armored articulated animals , to which the armadillos still existing today belong. Like these, the Pampatheria possessed an armor that covered the top of the head, the body and the tail. However, the pampatheria were significantly larger than the armadillos and reached a body weight of up to 200 kg. Despite their outward resemblance to the armadillos, the closest relatives are the gigantic Glyptodontidae . The Pampatheria most likely emerged during the Miocene , the oldest known finds are around 18 million years old. As with all secondary articulated animals, they originate in South America . In the course of the Pliocene - with the creation of a land bridge to North America - they also settled regions further north. At the end of the Pleistocene, the Pampatherien died out.
Compared to the glyptodonts and the armadillos, the pampatheria represent a rather poor group of forms with only a few representatives in the course of their tribal history. They mostly lived in more open and sometimes very dry landscapes and lived mainly on plants. The general physique speaks rather against a burrowing way of life, as it is proven in today's armadillos. However, it is possible that they continued to use individual burrows created by other graves. All in all, the Pampatherien are far less well researched than their relatives due to the rare finds. Representatives of the Pampatherien were first proven in the 1830s with finds from caves in southern Brazil. The family name that is valid today dates from 1954.
features
General and height
The representatives of the Pampatherien outwardly resembled today's armadillos (Dasypoda). They had a head with a slightly elongated snout and an armor that protected the skull, the body and the tail. However, they were significantly larger than the armadillos. In the late phase of their tribal history at the end of the Pleistocene , the largest forms such as Holmesina and Pampatherium reached a total length of 3 m and weighed a good 220 kg. A complete skeleton of Holmesina from the Gruta da Lapinha in eastern Brazil has a total length of 2.2 m. Earlier forms such as Vassallia were smaller, around 1 m long and weighing 60 to 90 kg, and were about the size of the giant armadillo, which is the largest member of today's armadillos. In comparison, this has a total length of 1.5 m and a weight of around 50 kg. Due to their size and their shell design, the Pampatheria are sometimes also called giant armadillos (" giant armadillos ").
Skull and dentition features

The skull of the Pampatherien resembles that of the armadillos, especially that of the euphractine forms with a closer relationship to the six-banded armadillo ( Euphractus sexinctus ), but was more massive, taller and wider. In large representatives it was between 28 and 36 cm long. The forehead line was often arched upwards , but there was a slight indentation in Kraglievichia . The rostrum was drawn out long, which differs significantly from the closely related Glyptodontidae with their greatly shortened skulls. As a result, the nasal bone also reached an enormous length of 14 to 16 cm. There was a severe necking of the skull behind the eye windows . The zygomatic arches bulged out far laterally. They had a sturdy structure with tall bones. In the side view, the anterior and posterior arch attachments were oriented strongly downwards, both met approximately at the level of the upper row of teeth. Here the surface was roughened and marks the starting point for strong muscles. There was a clearly pronounced crest on the parietal bones .
The lower jaw also had a massive structure and was significantly stronger than that of the armadillos. Ossification of the symphysis only occurred in very old individuals, otherwise the two halves of the lower jaw were usually not firmly attached. The lower jaw body showed a clearly downward curve on the lower edge, which is reminiscent of the glyptodonts. As a result, the lower jaw was also very high here. The ends of the joints reached very high. As with all articular animals, the dentition differed in its structural structure from that of most higher mammals . There were 9 teeth in each half of the jaw in both the upper and lower jaw, so the dentition consisted of a total of 36 teeth. All trained teeth as molarenartig ( molariform considered), cutting and canines , and premolars were not available. In contrast to the homodontic tooth design of the armadillos and the glyptodonts, the dentition in the pampatheria was heterodontic . The front teeth formed a short row and were shaped like nails with a rounded-oval cross-section. The posterior ones showed a clear longitudinal extension and had two transverse, flap-like widenings ( lobes ) at the respective ends , which gave the teeth a bilobatic character. The teeth lacked hard enamel , which is common practice for the secondary articular animals. Instead, they consisted of dentin , with predominantly hard orthodentin being formed inside . In some representatives, however, a rib made of particularly hard osteodentin ran along the central axis of the rear teeth on the chewing surface, which was thickened in the area of the lobes . However, the distribution of the hard components of the dentin varied greatly within the individual representatives of the pampatheria, which is interpreted as an adaptation to certain diets. In some representatives, the rear teeth were arranged in a step-like manner in the side view. The length of the row of teeth in the upper jaw reached up to 20 cm.
Body skeleton
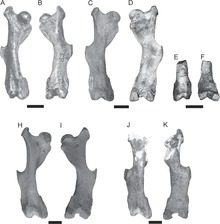
The spine of a complete individual was composed of 7 cervical, 10 thoracic, 1 lumbar, 11 sacrum and 22 tail vertebrae. Special features of the body skeleton are found primarily on the spine. As in the armadillos and the glyptodons, the cervical vertebrae were also fused together and formed a bony tube. This largely affects the axis (second cervical vertebra) and the following third to fifth cervical vertebrae, while the atlas (first cervical vertebra) was free to move. A special exception is Vassallia , a representative of the Pampatheria from the Upper Miocene , in which the atlas was also freely movable and not fused with the cervical tube. Additional articular surfaces ( xenarthric joints or xenarthrals) on the lateral processes of the posterior thoracic vertebrae and on the lumbar vertebrae are typical of all secondary articular animals . The musculoskeletal system showed only a few peculiarities. A very prominent third roll hillock (trochanter) appeared on the middle shaft of the thigh bone , which functioned as a muscle attachment point and is typical of secondary articulated animals; in the related Glyptodontidae this was clearly lower down near the knee. The tibia and fibula were fused together. The hands and feet were largely unspecialized, the inner and outer fingers of the hand were each reduced in length, but the three middle fingers were not clearly elongated as in the armadillos. All the rays had claws.
tank


Like all armored articulated animals, the pampatheria also had an outer, bony armor consisting of small bone platelets ( osteoderms ). The shell covered the top of the skull, the torso and the tail. In large exponents such as Holmesina, it was composed of over 850 individual elements, of which more than 630 are made up of body armor, over 50 on the head and around 130 on the tail. The body armor was similar to that of today's armadillos and consisted of a fixed shoulder and pelvis with movable straps in between. In contrast to the armadillos with their variable number of movable bands, the pampatheria always had three. The rigid and movable parts of the armor were made up of transverse rows of small bone platelets, which were probably covered with a horn.The bone platelets of the rigid armor had an elongated, mostly rectangular or polygonal shape and pointed on the surface in contrast to the sometimes complex pattern of the Armadillos and Glyptodons only had a central, flat elevation, which was surrounded by a horseshoe-shaped groove, with Vassallia and Pampatherium it appeared almost flat. The bone platelets were elongated on the movable ligaments and consisted of two sections, a short base part and a longer “tongue”. There were usually several deep holes on the surface. Due to the simple surface design, it is assumed for the pampatheria that only one horn plate covered an osteoderm and not several as in the armadillos, whose bone plates have a complex pattern on the surface. The tail was also completely surrounded by armor. The bone platelets here were arranged in rows like the armor, but unlike the armadillos, they did not overlap, but rather butted close together.
The osteoderms had a three-layer structure in cross-section. At the top and bottom there was a solid layer of bone, each enclosing an area that consisted of numerous cavities. It was about half the thickness of the bone plate. Sweat glands and hair follicles were embedded in this cancellous structure . The solid bone layer consisted alternately of hard layers and fiber bundles rich in collagen ( Sharpey's fibers ). A peculiarity of the bone platelets of the pampatheria was shown in the occurrence and the expression of these fiber bundles also on the side edges of the platelets of the stiff armor, where these had grown together with the neighboring platelets. In the armadillos, the Sharpey fibers have a different orientation in this area; in Glyptodont they have not yet been observed. Overall, the structure of the bone platelets gave them a certain elasticity and protected them from strong external forces.
distribution and habitat
Pampatheria were originally restricted to South America , where they developed at least since the Middle Miocene around 14 million years ago. They are found here mainly in the lowlands of the pamphlet region in the central area of the continent up to the Amazon basin , but in the latter their distribution was rather low. They were also found in the western coastal regions. In contrast to the related glyptodons and today's armadillos, there is no tradition from the high Andes . From the Middle Pliocene onwards , the Pampatheria also settled northern regions through the creation of the land bridge to North America . Fossil finds are mainly from the southern and southeastern areas of the USA from Texas over the Gulf Coast to Florida . Find sites very far north can be found in Kansas . The Pampatheria were adapted to more open landscapes and populated both the desert-like barren steppe areas of the Pampas and more humid coastal regions. Often, pampatheria and glyptodonts appear together in the fossil record . Both are not to be classified as ecologically identical, but only as similar.
Paleobiology
Locomotion
With regard to the musculoskeletal system, the pampatheria mediate between the smaller armadillos and the sometimes huge glyptodons. As with the Glyptodon, the hind legs are adapted to a clumsy gait. No major specializations are noticeable on the front legs. The upper articular process of the ulna, the olecranon , is comparatively shorter than that of the armadillos with their sometimes extremely long processes. A massive olecranon is often an indicator of a burrowing lifestyle, as is the case with most armadillos, as it increases the leverage of the triceps , which acts as the extensor for the forearm. The shape of the hand also speaks against significant digging activities, since the three middle fingers (II to IV) are not extended and no strong claws suitable for digging were developed. As active graves, armadillos often have strongly elongated middle fingers with strongly curved and elongated grave claws. The anatomical features of the pampatheria do not show any clear adaptation to an underground ( fossorial ) way of life.
However, tombs are known from the Miramar Formation in the area around Mar del Plata on the east coast of Argentina that date to the Lower and Middle Pleistocene . While larger structures between 0.9 and 1.8 m wide and sometimes up to 40 m in length can be traced back to large ground sloths from the group of Mylodontidae based on their size and the characteristic, two-line scratch marks , individual smaller ones are around 1 m wide and 75 cm high with other scratch marks. These generally have three lines with a maximum length of 30 cm. They correspond roughly to those of the front feet of the Pampatherien and could therefore be traced back to them. The size of the burrows corresponds roughly to that of the Pampatheria, whose shell could reach a diameter of 80 cm. They are also significantly larger than those of today's giant armadillo. In other, similarly large structures from southern Brazil, impressions of the osteoderms were also found on the walls, but these indicate rather large armadillos such as Propraopus . However, it cannot be ruled out that such burrows were used by other animals such as Pampatheria after they left the builder.
Diet
The teeth of the pampatheria are sometimes relatively high-crowned ( hypsodont ), which in comparison with other mammal groups rather indicates a preference for hard, abrasive grass food. Since the teeth lack enamel , it is not possible to examine the exact diet, for example in the form of isotope analyzes . Today's armadillos, which are equipped with pin-shaped teeth, include both omnivores and carnivores , and individual herbivores are also known among extinct forms . Special examinations of the chewing apparatus show that the pampatheria have similarities to today's ungulates with herbivorous diets. This includes, among other things, the high joint attachment on the lower jaw, which increased the leverage when chewing and thus allowed a higher biting force. This is also in relation to the clearly expansive zygomatic arches with their strongly roughened lower bony protuberance, which suggests a significantly stronger masticatory muscles , especially the masseter muscle attached here . Furthermore, the articular surface of the lower jaw is designed to be rather flat and thus supports horizontal chewing movements. This is often indicated by traces of abrasion running across the middle teeth. As with today's herbivores, the middle teeth are also greatly elongated and the entire row of teeth is clearly set back. On the rear molar-like teeth, harder dentin is also formed in the center of the occlusal surface, which increases the efficiency of the scissors when chewing. All these features, which can already be demonstrated in the earlier representatives such as Kraglievichia and Vassallia , indicate that the Pampatheria were adapted to plant food - possibly grasses. The main differences within the pampatheria are found in the development of the muscles, which is associated with the paleobiogeographical distribution of the individual forms and the resulting different food supply. For example, some species that were found in the more dry pamphlet region were adapted to much harder and more resistant plants, while those from the more humid coastal regions of today's Ecuador and Peru, which at that time were probably characterized by savannas or dry forests, preferred less abrasive plant food. In general, the pampatheria are seen as specializing in primarily grass-containing food with secondary, landscape-dependent adaptation to firmer or fibrous parts of the plant. It is also noteworthy that the Pampatherien have a relatively large chewing surface of all teeth in relation to their body size, which is much larger than the closely related Glyptodontidae . With a body weight of 177 kg , Holmesina had a chewing surface of around 1520 mm², which is just below today's tapirs, which have a body weight of 210 to 290 kg between 1530 and 2240 mm². In comparison, the glyptodont representative Plohophorus , who weighed around 260 kg, had only almost half of the chewing surface with 900 mm². It is possible that the less complex tooth design of the pampatheria compared to the glyptodonts and the reduction in sharp-edged ribs on the tooth surfaces led to an enlargement of the chewing surface and thus a better utilization of the ingested food in the oral cavity.
brain
A complete pouring of the brain capsule from Pampatherium could be examined. This comprised a volume of around 133 cm³ with an assumed body weight of the animal of 210 kg. The encephalization quotient was thus 0.27. A value of 0.17 is assumed for the older Vassallia . Both are within the range of the Glyptodontidae (0.12 to 0.4), but in the lower range of the armadillos (0.44 to 1.06). Comparable to the glyptodons and armadillos, the olfactory bulb was extremely large and took up about 8% of the total volume of the brain. Two-thirds also claimed the cerebrum , the rest fell to the cerebellum . The olfactory bulb had a narrow-pointed shape, which corresponded to that of the glyptodons. In contrast, the armadillos have a rather broad olfactory bulb. Only the sulcus suprasylvianus of the parietal lobe was recognizable in the relatively simple relief of the cerebral cortex , while the additionally developed sulcus praesylvianus in the armadillos was missing.
Systematics
External system
|
Relationship between pampatheria and armadillos according to morphological data from Billet et al. 2011
|

The Pampatheriidae represent a family within the order of the armored secondary animals (Cingulata). This order, which was once rich in forms in South and North America, also includes the also extinct Glyptodontidae , Peltephilidae , the Pachyarmatheriidae and Palaeopeltidae as well as the armadillos (Dasypoda) that still exist today . The armored articulated animals are in turn included in the superordinate order of the articulated animals (Xenarthra), which also includes the tooth arms (Pilosa) with today's sloths (Folivora) and the anteaters (Vermilingua). The secondary articulated animals represent one of the four main lines of the higher mammals , which stands opposite the other three (collectively as Epitheria ) as a sister group . A general common feature of the secondary articular animals consists in the named xenarthric joints (secondary joints, also xenarthrals) on the articular processes of the posterior thoracic and lumbar vertebrae. The Glyptodontidae, whose lumbar vertebrae are fused together as a unique feature, are an exception . The origin of the articular animals has not yet been clarified, the oldest fossil finds that are counted among the armadillos come from South America and belong to the Paleocene more than 56 million years ago. Based on molecular genetic studies, it is probable that the secondary animals split off from the other higher mammals as early as the end of the Lower Cretaceous around 103 million years ago. The armadillos as the closest relatives of the Pampatherien living today separated from the common line with the tooth arms at the beginning of the Paleocene about 65 million years ago.
The closest relatives of the Pampatherien within the armored collateral animals are the glyptodons. These are characterized by a completely rigid back armor and also reached an enormous size with a body weight of up to 2 t. Due to the characteristic construction of the armor with the three movable bands, the Pampatheria were originally thought to be more closely related to the armadillos; they were run within this in their own subfamily. However, studies on skulls and teeth showed a closer relationship between the pampatheria and the glyptodonts. Among other things, this could be proven in the structure of the ear canal and in the construction of the chewing apparatus, such as the high lower jaw. In addition, the teeth of the representatives of both families are more complex. The armadillos have simple, nail-like teeth, the pampatheria and glyptodonts these are clearly elongated and characterized by two or three transverse lobes . The Pampatherien and Glyptodonten together form the superordinate group of the Glyptodonta . The Pachyarmatheriidae proved to be a sister group of the Glyptodonta, whose characteristic is an overlapping back armor without movable ligaments in between. They were widespread in South and North America from the Upper Miocene to the beginning of the Holocene . According to skeletal anatomical studies from 2006 and 2011, the Glyptodonta themselves are embedded more deeply in the group of armadillos, as they may have a closer connection to the Euphractinae , i.e. the group around the six-banded armadillo , the dwarf armadillo and the bristle armadillos . Together with these, they form an evolutionary line of development. In contrast, in 2016 published showed in molecular genetic studies of doedicurus from the group of Glyptodonten a closer relationship with the Tolypeutinae ( armadillos , cabassous and giant armadillo ) and Chlamyphorinae (Gürteltmulle). To what extent the pampatheria occupy a position similar to that of the glyptodonts is unclear, as no genetic material has yet been obtained. In both research results, i.e. both the skeletal-anatomical and the molecular-genetic, the armadillos form a paraphyletic group. The other groups such as the Palaeopeltidae and Peltephilidae are in a more or less close relationship to the other armored articulated animals.
Internal system
The Pampatheria are considered to be a rather conservative group that has barely undergone any development and therefore appears less varied than their close relatives, the armadillos and glyptodons. Subfamilies are not differentiated within the Pampatherien family. In general, the genera and species are identified in a similar way to the other groups of armored articulated animals on the basis of the different formation of the osteoderms of the armor and not, as is usual in mammals, with the help of skull and skeletal features. The reason for this is the relatively high frequency of bone platelets compared with skeletal material. Based on the bone platelets, two lines of development can be identified within the group. One includes Vassallia and Pampatherium , both of which are characterized by little ornamented armor plates, the other can be assumed to be Plaina , Kraglievichia and Holmesina .
Overview of the genera
About half a dozen genera are recognized:
- Family: Pampatheriidae (= Chlamydotheriidae) Paula Couto , 1954
- Scirrotherium Edmund & Theodor , 1997
- Kraglievichia Castellanos , 1927
- Vassallia Castellanos , 1927
- Plaina Castellanos , 1937
- Tonnicinctus Góis, González Ruiz, Scillato-Yané & Soibelzon , 2015
- Pampatherium (= Chlamytherium , Chlamydotherium , Hoffstetteria ) Gervais & Ameghino , 1880
- Holmesina Simpson , 1930
The following genus is considered unsafe to belong to the pampatheria:
- Machlydotherium Ameghino , 1902
The delimitation of the individual genera is assessed differently. Some researchers also see Plaina as synonymous with Kraglievichia . In the past, Holmesina was seen as being partially identical to Pampatherium . In general, a revision of the family is requested.
Tribal history
Origins
The origin of the Pampatherien, like that of the other collateral animals, is most likely in South America . The oldest representative is partly Machyldotherium from the Upper Eocene 35 to 37 million years ago. Finds come from Gran Barranca south of Lake Colhué Huapi in northern Patagonia . Other fossil remains in the form of bone platelets come from the somewhat younger Sarmiento Formation in Barrancas Blancas , northeast of Lake Colhué-Huapi , also Patagonia, as well as from the Guabirotuba Formation in the southern Brazilian state of Paraná . The position of Machyldotherium within the Pampatheria is controversial.
Miocene

By the uncertain allocation of Machyldotherium the described only in 1997 applies Scirrotherium than previously undoubtedly the oldest member of the line of Pampatherien. One of the earliest records is a bone plate from the Castillo Formation in northwestern Venezuela , the age of which dates from 19 to 17 million years and thus to the Lower Miocene . More extensive material comes from the La Venta formation in Colombia and consists of a skull only 14 cm long with a lower jaw and also numerous remains of the body skeleton and some bone plates. Their age is around 14 to 15 million years, which corresponds to the Middle Miocene. The finds point to a very small relative with a body weight of only about 40 kg. Only at the beginning of the Upper Miocene is the genus also documented for the southern areas of South America. These include finds from the Paraná Formation in the Argentine province of Entre Ríos and the Puerto Madryn Formation in the extreme northeast of the Argentine province of Chubut . Both rock units are actually characterized by marine sediments , including scirrotherium bone platelets and individual terrestrial elements. The finds are estimated to be around 8.7 million years old. Outstanding is the very fossil-rich Conglomerodo ossifero of the Ituzaingó Formation , which is opened up by the Río Paraná in Entre Ríos. The extensive remains of the secondary articulated animals also contain numerous osteoderms of Scirrotherium , which has its earliest evidence in research history outside of northern South America. In addition, skull remains and parts of the body skeleton of Kraglievichia , another early and relatively small representative , could also be discovered here. Vassallia is also one of these early forms , which was described using a lower jaw and a few bone plates from the Andalhuala formation in the Argentine province of Catamarca . A nearly complete skull from the Corral Quemado Formation in the same province can also be assigned to this genus .
Even before the onset of the Great American Faun Exchange , which was triggered by the creation of the land bridge between South and North America with the closure of the Isthmus of Panama during the Pliocene , Pampatheria can be found in the Upper Miocene north of the South American continent. The Curré formation in southern Costa Rica comes from bone platelets and remains of the body skeleton of Scirrotherium . It is possible that before the collision of the two continental masses there were smaller islands or arches of islands that were settled by the Pampatheria.
Pliocene and Pleistocene


The Pliocene is mainly characterized by the further expansion of the Pampatheria to the north. From deposits in the basin of San Miguel Allende in the central Mexican state of Guanajuato , osteoderms have been passed down that are dated 4.7 to 4.8 million years ago and belong to the genus Plaina . In the subsequent Pleistocene , only the terminal forms Holmesina , Pampatherium and Tonnicinctus have survived, but the latter has so far been limited to only two sites.
Holmesina first appeared about 2.0 to 2.6 million years ago at the transition from the Pliocene to the Pleistocene in North America. It is believed that the genus there originated from previous immigrants such as Plaina . One of the oldest finds is a nearly complete skeleton from the important Haile fossil site , 6 km northeast of Newberry in Alachua County , Florida . The finds came to light more precisely in the location 15A, a sediment-filled limestone fissure. The first immigrants in North America were still relatively small, they weighed around 44 kg with a thigh bone length of around 20 cm. By the end of the Pleistocene, they gained many times their body weight and then weighed around 220 kg. These large forms then had thigh bones with a length of 33 cm. Holmesina also returned to South America during the Pleistocene, where it is mainly recorded in the Upper Pleistocene. Here it evidently went through a new differentiation and is therefore passed down with several species. Among other things , two complete skeletons have been recovered from the Gruta da Lapinha near Iramaia in the Brazilian state of Bahia . When exactly this return migration started is unknown; in El Salvador remains of Holmesina in the form of bone platelets are already known from the transition from the Old to the Middle Pleistocene.
For Pampatherium , however, it is assumed that it developed in the Lower Pleistocene in South America from forms such as Vassallia . Similar to Holmesina, it experienced a massive increase in size towards the end of the Pleistocene. It may also have reached Central and North America in another wave of emigration. It did not advance as far north as its relative, however; the northernmost finds come from the Upper Pleistocene of Mexico , for example from Térapa. Reasons for the more limited spread of Pampatherium could lie in the better adaptation to clearly drier climatic conditions of the genus.
die out
Both Pampatherium and Holmesina died out at the end of the Pleistocene during the Quaternary wave of extinction . Young data for Pampatherium comes from the Brazilian state of Bahia and is around 13,000 BP . Radiocarbon dates for Holmesina from La Brea in northwestern Peru are only slightly older, at 13,600 to 14,400 BP . From the Cupisnique desert further south on the Peruvian west coast, from where relatively extensive skeletal material from Holmesina originates, there are age data that vary between 8,900 and 25,000 BP. The Quaternary extinction wave in America coincides with the initial colonization of both continents by early hunters and gatherers , which began about 15,000 years ago. It is unclear whether active hunting led to the disappearance of the pampatheria. In North America, of around two dozen sites from the end of the Pleistocene that contain the remains of Pampatheria, only a few are known to be associated with early human hunting. This includes Lubbock Lake , Texas, a major, 120 ha large reference in an old arm of the Yellow House Draw, a tributary of the Brazos River . The bottom layer of the find is dated 11,100 years ago and, in addition to numerous bone fragments, also contained stone tools , especially hammer stones and stone working documents (anvils). The bones, including Holmesina , show punch marks and thus indicate that they were used as a food source. The finds from Holmesina at Lubbock Lake are among the youngest known of the Pampatheria.
Research history

The exploration of the Pampatherien goes back to the first half of the 19th century. This makes them one of the first animals that were previously endemic in America and known to science. In 1836, the Danish paleontologist Peter Wilhelm Lund (1801–1880) discovered the remains of pampatheria in caves near Lagoa Santa in the Brazilian state of Minas Gerais and published two preliminary reports in November 1837, where he also mentioned the first finds of articulated animals . A more extensive treatise was put on paper in 1839, but it was only to appear another two years later. In an overview article on Lund's research that HC Örsted has compiled in the meantime, he mentioned the new genus Chlamytherium . Lund's own report was then published in 1841. In this he presented the genus Chlamydotherium , the material that was available to him for his description included some bone platelets and jaw fragments. (As early as 1839, brief reports on Lund's work, which he had sent by letter and in which he also mentions Chlamydotherium as an armadillo the size of a tapir , appeared in several important scientific journals in Europe, for example in the editions of the London Natural History Museum or the Paris Muséum national d'histoire naturelle .) The generic name Chlamydotherium is made up of the Greek words χλαμύς ( chlamys "coat", genitive χλαμύδος chlamydos ) and θηρίον ( thēríon "animal"). However, already in 1838 Heinrich Georg Bronn had given a representative of the Glyptodontidae , close relatives of the Pampatheria, the name Chlamydotherium based on finds from Brazil and Uruguay , whereby it was preoccupied (but today it is synonymous with Glyptodon ).
In the second half of the 19th century, little was known about extinct armadillos and pampatheria, in contrast to the glyptodons. In 1875, the Argentine researcher Florentino Ameghino (1854–1911) first mentioned Pampatherium in a manuscript , probably in ignorance of Lund's Chlamytherium or Chlamydotherium . However, his publication has not survived, and possibly never appeared. However, together with Henri Gervais , he realized in 1880 that Pampatherium is identical to Lund's Chlamydotherium (both authors do not recognize at this time that the name was preoccupied by a representative of the Glyptodon). Due to the structure of the carapace with flexible bands, they emphasize the close connection between Chlamydotherium and the armadillos, but also notice similarities with the glyptodons due to the structure of the teeth of the lower jaw. Chlamytherium is not mentioned in the scriptures, in the following years until well into the 20th century it was often used as a generic name.
The first bone platelet finds in North America were discovered by Joseph Willcox in 1888 at Peace Creek in Florida . He passed this on to the American paleontologist Joseph Leidy (1823-1891), who described it the following year as belonging to Glyptodon . In the same year, after he recognized the similarities in the design of the bone platelets, Leidy corrected the genus belonging to Lund's Chlamytherium . Further fossil remains also followed from Florida and Texas and included not only the osteoderms but also the lower jaw and parts of the skull. These showed that there are some differences to the South American forms. In 1930, George Gaylord Simpson (1902-1984) named the genus Holmesina , citing a lower jaw and individual upper jaw fragments from Bradenton Field in Florida .
Ameghino had already coined the family name Chlamydotheriidae in 1889, which is based on his finding that Pampatherium is only a synonym for Lund's Chlamydotherium . In 1930 Oliver Perry Hay changed this to Chlamytheriinae, in addition to correcting the name, moving the group to the level of the subfamily within the armadillos. Carlos de Paula Couto (1910-1982) then pointed out in 1954 that the name Pampatherium used by Ameghino in 1875 is valid and is preferable to Chlamytherium , since the latter name became invalid at the moment when Lund decided to rename it Chlamydotherium . His then proposed name Pampatheriidae for the family slowly established itself in the period that followed.
Individual evidence
- ^ Gordon Edmund: The fossil giant armadillos of North America (Pampatheriinae, Xenarthra Edentata). In: G. Gene Montgomery (Ed.): The Ecology and Evolution of Armadillos, Sloths, and Vermilinguas. Smithsonian Institution Press, 1985, pp. 83-93
- ↑ a b c Timothy J. Gaudin and John R. Wible: The Phylogeny of Living and Extinct Armadillos (Mammalia, Xenarthra, Cingulata): A Craniodental Analysis. In: MT Carrano, TJ Gaudin, RW Blob and JR Wible (eds.): Amniote Paleobiology. Chicago / London: University of Chicago Press, 2006, pp. 153-198
- ^ A b H. Gregory McDonald: Paleoecology of extinct Xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45 (4), 2005, pp. 313-333
- ↑ a b c Jorge Felipe Moura, Flávio Góis, Fernando Carlos Galliari and Marcelo Adorna Fernandes: A new and most complete pampathere (Mammalia, Xenarthra, Cingulata) from the Quaternary of Bahia, Brazil. Zootaxa 4661 (3), 2019, pp. 401-444, doi: 10.11646 / zootaxa.4661.3.1
- ↑ In it A. Croft and Velizar Simeonovski: Horned armadillos and rafting monkeys. The fascinating fossil mammals of South America. Indiana University Press, 2016, pp. 1–304 (pp. 196–197)
- ↑ Arnaud Leonard Jean Desbiez and Danilo Kluyber: The Role of Giant Armadillos (Priodontes maximus) as Physical Ecosystem Engineers. Biotropica 45 (5), 2013, pp. 537-540
- ↑ a b Jim I. Mead, Sandra L. Swift, Richard S. White, H. Greg McDonald and Arturo Baez: Late Pleistocene (Rancholabrean) Glyptodont and Pampathere (Xenarthra, Cingulata) from Sonora, Mexico. Revista Mexicana de Ciencias Geológica 24 (3), 2007, pp. 439-449
- ↑ a b c d e f g h G. J. Scillato-Yané, AA Carlini, EP Tonni and JI Noriega: Paleobiogeography of the late Pleistocene pampatheres of South America. Journal of South American Earth Sciences 20, 2005, pp. 131-138
- ↑ a b c d Flávio Góis, Gustavo Juan Scillato-Yané, Alfredo Armando Carlini and Martín Ubilla: Una nueva especie de Holmesina Simpson (Xenarthra, Cingulata, Pampatheriidae) del Pleistoceno de Rondônia, Sudoeste de la Amazonia, Brazil. Revista Brasileira de Paleontologia 15 (2), 2012, pp. 211-227
- ↑ a b c Gerardo De luliis, María S. Bargo and Sergio F. Vizcaíno: Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. Journal of Vertebrate Paleontology 20 (4), 2000, pp. 743-754
- ↑ a b c Sergio F. Vizcaíno, Gerardo De luliis and María S. Bargo: Skull shape, masticatory apparatus, and diet of Vassalliaand Holmesina (Mammalia: Xenarthra: Pampatheriidae). When anatomy constrains destiny. Journal of Mammalian Evolution 5 (4), 1998, pp. 291-322
- ↑ Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). Paleobiology 35 (3), 2009, pp. 343-366
- ^ Daniela C. Kalthoff: Microstructure of Dental Hard Tissues in Fossil and Recent Xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology 272, 2011, pp. 641-661
- ↑ a b c d Gerardo De Iuliis and A. Gordon Edmund: Vassallia maxima Castellanos, 1946 (Mammalia: Xenarthra: Pampatheriidae), from Puerta del Corral Quemado (Late Miocene to Early Pliocene), Catamarca Province, Argentina. Robert J. Emry (Ed.): Cenozoic mammals of land and sea: Tributes to the Career of Clayton E. Ray. Smithonia Contributions to Paleobiology 93, 2002, pp. 49-63
- ↑ Timothy J. Gaudin: The Morphology of Xenarthrous Vertebrae (Mammalia: Xenarthra). Fieldiana Geology, New Series 41, 1999, pp. 1-38
- ↑ H. Gregory McDonald: Xenarthran skeletal anatomy: primitive or derived? Senckenbergiana biologica 83, 2003, pp. 5-17
- ↑ a b N. Milne, Sergio. F. Vizcaíno and JC Fernicola: A 3D geometric morphometric analysis of digging ability in the extant and fossil cingulate humerus. Journal of Zoology 278 (1), 2009, pp. 48-56
- ↑ a b Gordon Edmund: The Armor of the fossil giant armadillos (pampatheriidae, Xenarthra, Mammalia). Texas Memorial Museum, The University of Texas, Austin, 1985, pp. 1-20
- ↑ Santiago Rodriguez-Bualó, Alfredo Eduardo Zurita, Flávio Góis, Ángel R. Miño-Boilini, Esteban Soibelzon and Freddy Paredes-Ríos: Pampatheriidae (Xenarthra, Cingulata) from the Tarija valley, Bolivia: A taxonomic update. Rivista Italiana di Palrontologia e Stratigrafia 120 (2), 2014, pp. 253-259
- ^ A b Gideon T. James: An Edentate from the Pleistocene of Texas. Journal of Paleontology 31 (4), 1957, pp. 796-808
- ^ Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. Journal of Morphology 267, 2005, pp. 1441-1460
- ↑ Dominik Wolf, Daniela C. Kalthoff and P. Martin Sander: Osteoderm Histology of the Pampatheriidae (Cingulata, Xenarthra, Mammalia): Implications for Systematics, Osteoderm Growth, and Biomechanical Adaptation. Journal of Morphology 273, 20912, pp. 388-404
- ^ A b François Pujos and Rodolfo Salas: A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de l'Institut Français d'Etudes Andines 33 (2), 2004, pp. 331-377
- ^ Claude W. Hibbard, Richard J. Zakrzewski, Ralph E. Eshelman, Gordon Edmund, Clayton D. Griggs, and Caroline Griggs: Mammals from the Kanopolis Local Fauna, Pleistocene (Yarmouth) of Ellsworth County, Kansas. Museum of Paleontology, University of Michigan 25 (2), 1978, pp. 11-48
- ^ David D. Gillette and Clayton E. Ray: Glyptodonts of North America. Smithonian Contributions to Paleobiology 40, 1981, pp. 1-251
- ↑ a b Sergio F. Vizcaíno, Marcelo Zaráte, M. Susana Bargo and Alejandro Dondas: Pleistocene burrows in the Mar del Plata area (Argentina) and their probable builders. Acta Palaeontologica Polonica 46 (2), 2001, pp. 289-301
- ↑ Alejandro Dondas, Federico I. Isla and José L. Carballido: Paleocaves exhumed from the Miramar Formation (Ensenadan Stage-age, Pleistocene), Mar del Plata, Argentina. Quaternary International 210, 2009, pp. 44-50
- ^ Francisco Sekiguchi Buchmann, Renato Pereira Lopes and Felipe Caron: Paleotoca do Município de Cristal, RS. Registro da atividade fossorial de mamíferos gigantes extintos no sul do Brasil. In: M. Winge, C. Schobbenhaus, CRG Souza, ACS Fernandes, M. Berbert-Born, W. Sallun filho and ET Queiroz: (Eds.): Sítios Geológicos e Paleontológicos do Brasil. 2010, pp. 1–10
- ^ Sergio F. Vizcaíno, Richard A. Fariña, M. Susana Bargo and Gerardo De Iuliis: Functional and phylogenetic assessment of the masticatory adaptations in Cingulata (Mammalia, Xenarthra). Ameghiniana 41 (4), 2004, pp. 651-664
- ↑ Sergio F. Vizcaíno, M. Susana Bargo and Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. Ameghiniana 43 (1), 2006, pp. 11-26
- ↑ P. Sebastián Tambusso and Richard A. Fariña: Digital cranial endocast of Pseudoplohophorus absolutus (Xenarthra, Cingulata) and its systematic and evolutionary implications. Journal of Vertebrate Paleontology 35 (5), 2015, p. E967853, doi: 10.1080 / 02724634.2015.967853
- ↑ P. Sebastián Tambusso and Richard A. Fariña: Digital endocranial cast of Pampatherium humboldtii (Xenarthra, Cingulata) from the Late Pleistocene of Uruguay. Swiss Journal of Palaeontology 134, 2015, pp. 109–116, doi: DOI 10.1007 / s13358-015-0070-5
- ↑ a b c Guillaume Billet, Lionel Hautier, Christian de Muizon and Xavier Valentin: Oldest cingulate skulls provide congruence between morphological and molecular scenarios of armadillo evolution. Proceedings of the Royal Society B, 278, 2011, pp. 2791-2797
- ↑ Maureen A. O'Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R Wible and Andrea L. Cirranello: The Placental Mammal Ancestor and the Post-K-Pg Radiation of Placentals. Science 339, 2013, pp. 662-667, doi: 10.1126 / science.1229237
- ↑ Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, pp. 1–431 (pp. 200–204)
- ^ Sergio F. Vizcaíno and WJ Loughry: Xenarthran biology: Past, present and future. In: Sergio F. Vizcaíno and WJ Loughry (eds.): The Biology of the Xenarthra. University Press of Florida, 2008, pp. 1-7.
- ↑ Frédéric Delsuc, Michael J. Stanhope and Emmanuel JP Douzery: Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyzes of mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 28, 2003, pp. 261-275
- ↑ Frédéric Delsuc, Sergio F Vizcaíno and Emmanuel JP Douzery: Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. BMC Evolutionary Biology 4 (11), 2004, pp. 1-13
- ↑ Leopold Héctor Soibelzon, Martín Zamorano and Gustavo Juan Scillato-Yané: Un Glyptodontidae de gran tamaño en el Holoceno Temprano de la Region Pampeana, Argentina. Revista Brasileira de Paleontologia 15 (1), 2012, pp. 105-112
- ↑ Juan-Carlos Fernicola, Serdio F. Vizcaíno and Richard a. Fariña: The evolution of armored xenarthrans and the phylogeny of the glyptodonts. In: Sergio F. Vizcaíno and WJ Loughry (eds.): The Biology of the Xenarthra. University Press of Florida, 2008, pp. 79-85
- ↑ Juan C. Fernicola, Andrés Rinderknecht, Washington Jones, Sergio F. Vizcaíno and Kleberson Porpino: A new species of Neoglyptatelus (Mammalia, Xenarthra, Cingulata) from the late Miocene of Uruguay provides new insights on the evolution of the dorsal armor in cingulates . Ameghiniana 55, 2018, pp. 233-252
- ↑ Frédéric Delsuc, Gillian C. Gibb, Melanie Kuch, Guillaume Billet, Lionel Hautier, John Southon, Jean-Marie Rouillard, Juan Carlos Fernicola, Sergio F. Vizcaíno, Ross DE MacPhee and Hendrik N. Poinar: The phylogenetic affinities of the extinct glyptodonts. Current Biology 26, 2016, pp. R155-R156, doi: 10.1016 / j.cub.2016.01.039
- Jump up ↑ Kieren J. Mitchell, Agustin Scanferla, Esteban Soibelzon, Ricardo Bonini, Javier Ochoa and Alan Cooper: Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos. Molecular Ecology 25 (14), 2016, pp. 3499-3508, doi: 10.1111 / mec.13695
- ↑ Daniel Perera, Pablo Martín and Torino Ciancio: La Presencia del Xenartro Palaeopeltis inornatus Ameghino, 1894, en la Formación Fray Bentos (Oligoceno Tardío), Uruguay. Ameghiniana 51 (3), 2014, pp. 254-258
- ↑ a b Richard M. Fariña, Sergio F. Vizcaíno and Gerardo de Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, pp. 1-436 (pp. 220-223) ISBN 978-0-253-00230-3
- ↑ a b Malcolm C. McKenna and Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, pp. 1-631 (pp. 86-87)
- ↑ a b c Flávio Góis, Gustavo Juan Scillato-Yané, Alfredo Armando Carlini and Edson Guilherme: A new species of Scirrotherium Edmund & Theodor, 1997 (Xenarthra, Cingulata, Pampatheriidae) from the late Miocene of South America. Alcheringa: An Australasian Journal of Palaeontology 37 (2), 2013, pp. 177-188
- ↑ a b Flávio Góis, Laureano Raúl González Ruiz, Gustavo Juan Scillato-Yané and Esteban Soibelzon: A Peculiar New Pampatheriidae (Mammalia: Xenarthra: Cingulata) from the Pleistocene of Argentina and Comments on Pampatheriidae Diversity. PlosOne 10 (6), 2015, p. E0128296 doi : 10.1371 / journal.pone.0128296
- ↑ a b c Castor Cartelle Guerra and Germán Arturio Bohórquez Mahecha: Pampatherium paulacoutoi, uma nova espécie de tatu gigante da Bahia, Brasil (Edentata, Dasypodidae) . Revista Brasileira de Zoologia, 2, 1984, pp. 229-254
- ↑ Richard F. Kay, Richard H. Madden, M. Guiomar Vucetich, Alfredo A. Carlini, Mario M. Mazzoni, Guillermo H. Re, Matthew Heizleri and Hamish Sandeman: Revised geochronology of the Casamayoran South American Land Mammal Age: Climatic and biotic implications. PNAS 96 (23), 1999, pp. 13235-13240
- ^ Alfredo A. Carlini, M. Ciancio and Gustavo J. Scillato-Yané: Los Xenarthra de Gran Barranca, more de 20 Ma de historia. Congreso Geológico Argentino 16 (4), 2005, pp. 419-424
- ↑ María Teresa Dozo, Martín Ciancio, Pablo Bouza and Gastón Martínez: Nueva asociación de mamíferos del Paleógeno en el este de la Patagonia (provincia de Chubut, Argentina): implicancias biocronológicas y paleobiogeográficas. Andean Geology 41 (1), 2014, pp. 224-247, doi : 10.5027 / andgeoV41n1-a09 .
- ↑ Fernando A. Sedor, Édison V. Oliveira, David D. Silva, Luiz A. Fernandes, Renata F. Cunha, Ana M. Ribeiro and Eliseu V. Dias: A New South American Paleogene Land Mammal Fauna, Guabirotuba Formation (Southern Brazil ). In: Journal of Mammalian Evolution 24, 2017, pp. 39-55
- ^ Robert Hoffstetter: Remarques sur la phylogénie et la classification des édentés xénarthres (mammifères) actuels et fossiles. Bulletin du Muséum National d'Histoire Naturelle, 2nd série 41 (1), 1969, pp. 91-103
- ^ Ascanio D. Rincón, Andrés Solórzano and H. Gregory McDonald: New mammal records from the Early Miocene Castillo Formation of Venezuela. Journal of Vertebrate Paleontology 28 (3; suppl), 2010, p. 151a
- ↑ Ascanio D. Rincón, Andrés Solórzano, Mouloud Benammi, Patrick Vignaud and H. Gregory McDonald: Chronology and geology of an Early Miocene mammalian assemblage in North of South America, from Cerro La Cruz (Castillo Formation), Lara State, Venezuela: implications in the 'changing course of Orinoco River' hypothesis. Andean Geology 41 (3), 2014, pp. 507-528, doi : 10.5027 / andgeoV41n3-a02 .
- ^ A. Gordon Edmund and Jessica Theodor: A new giant pampatheriid armadillo. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli, and John J. Flynn (Eds.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, pp. 227-232
- ^ Alfredo A. Carlini, Sergio F. Vizcaíno and Gustavo J. Scillato-Yané: Armored Xenarthtans: A unique taxonomic and ecologic assemblage. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli, and John J. Flynn (Eds.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, pp. 213-226
- ^ Adriana M. Candela, Ricardo A. Bonini and Jorge I. Noriega: First continental vertebrates from the marine Parana 'Formation (Late Miocene, Mesopotamia, Argentina): Chronology, biogeography, and paleoenvironments. Geobios 45, 2012, pp. 515-526
- ↑ María Teresa Dozo, Pablo Bouza, Alejandro Monti, Luis Palazzesi, Viviana Barreda, Gabriela Massaferro, Roberto A. Scasso and Claudia P. Tambussi: Late Miocene continental biota in Northeastern Patagonia (Península Valdés, Chubut, Argentina). Palaeogeography, Palaeoclimatology, Palaeoecology 297, 2010, pp. 100-109
- ↑ Alberto Luis Cione, María de las Mercedes Azpelicueta, Mariano Bond, Alfredo A. Carlini, Jorge R. Casciotta, Mario Alberto Cozzuol, Marcelo de la Fuente, Zulma Gasparini, Francisco J. Goin, Jorge Noriega, Gustavo J. Scillato-Yané , Leopoldo Soibelzon, Eduardo Pedro Tonni, Diego Verzi and María Guiomar Vucetich: Miocene vertebrates from Entre Ríos province, eastern Argentina. In: FG Aceñolaza and R. Herbst (eds.): El Neógeno de Argentina. Universidad Nacional de Tucumán, 2000, pp. 191-237
- ↑ Gustavo J. Scillato-Yané, Flávio Góis, Alfredo E. Zurita, Alfredo A. Carlini Laureano R. Gonzáles Ruiz, Cecilia M. Krmpotic, Cristian Oliva and Martín Zamorano: Los Cingulata (Mammalia, Xenarthra) del “Conglomerado osífero” ( Mioceno Tardío) de la Formación Ituzaingó de Entre Ríos, Argentina. In: Diego Brandoni and Jorge I. Noriega (eds.): El Neógeno de la Mesopotamia argentina. Buenos Aires, 2013, pp. 118-134
- ^ César A. Laurito and Ana L. Valerio: Scirrotherium Antelucanus, una nueva especie de Pampatheriidae (Mammalia, Xenarthra, Cingulata) del Mioceno Superior de Costa Rica, América Central. - Revista Geológica de América Central 49, 2013, pp. 45-62
- ↑ Oscar Carranza Castañeda and Wade E. Miller: Late Tertiary terrestrial mammals from Central Mexico and their relationship to South American immigrants. Revista Brasileira de Paleontologia 7 (2), 2004, pp. 249-261
- ↑ Michael O. Woodburne: The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens. Journal of Mammal Evolution 17, 2010, pp. 245-264
- ↑ a b Jesse Steadman Robertson: Latest Pliocene mammals from Haile 15A, Alachua County, Florida. Bulletin of the Florida State Museum, Biological Sciences 20, 1976, pp. 111-186 (pp. 124-142)
- ^ Daniel H. Aguilar and César A. Laurito: El armadillo gigante (Mammalia, Xenarthra, Pampatheriidae) del Río Tomayate, Blancano tardío-Irvingtoniano temprano, El Salvador, América Central. Revista Geológica de América Central 41, 2009, pp. 25-36
- ^ Donald K. Grayson and David J. Meltzer: Clovis Hunting and Large Mammal Extinction: A Critical Review of the Evidence. Journal of World Prehistory 16 (4), 2002, pp. 313-359
- ^ J. Tyler Faith and Todd A. Surovell: Synchronous extinction of North America's Pleistocene mammals. PNAS 106, 2009, pp. 20641-20645
- ↑ Eileen Johnson and Vance T. Holliday: Lubbock Lake: Late Quaternary cultural and environmental change on the Southern High Plains, USA. Journal of Quaternary Science 4 (2), 1989, pp. 145-165
- ^ Vance T. Holliday: Late Pleistocene and Holocene stratigraphy, Southern High Plains of Texas. Geological Society of America, Centennial Field Guide - South-Central Section 1988, pp. 293-298
- ↑ Peter Wilhelm Lund: Om Huler i Kalksteen i det indre af Brazil, the tildeels indeholde fossil knokler. Andean afhandling. Lapa de Cerca Grande. Det Kongelige Danske Videnskabernes Selskbas Naturvidenskabelige og Matematiske Afhandlinger 6, 1837, pp. 207–249 and 309–332 ( [1] )
- ↑ Peter Wilhelm Lund: Blik paa Brazil Dyreverden för Sidste Jordomvaeltning. Andean Afhandling: Patteedyrene. Det Kongelige Danske Videnskabernes Selskbas Naturvidenskabelige og Matematiske Afhandlinger 8, 1839, pp. 61–144 (p. 69) ( [2] )
- ^ Peter Wilhelm Lund: List of fossil Mammifera from the Basin of Rio das Velhas, with an extract of some of their distinguishing characters. Annals of Natural History 3, 1839, pp. 422–427 ( [3] )
- ^ Peter Wilhelm Lund: Coup d'oeil sur les espèces éteintes de mammifères du Brésil; Extrait de quelques mémoires presentés à l'Académie Royale des Sciences de Copenhague. Annales des sciences naturelles, Zoologie 2 (11), 1939, pp. 214–234 ( [4] )
- ↑ Heinrich Georg Bronn: Lethaea geognostica: or, illustrations and description of the most characteristic fossils for the mountain formations, Volume 2. Stuttgart, 1838, pp. 1–1346 (pp. 1256–1259) ( [5] )
- ^ Cástor Cartelle: Peter W. Lund, a naturalist of several sciences. Lundiana 3 (2), 2002, pp. 83-85
- ^ Henri Gervais and Florentino Ameghino: Les mammifères fossiles de l´Ámérique du Sud. Paris-Buenos Aires, 1880, pp. 1–225 (pp. 210–211)
- ↑ Alvin R. Cahn: Chlamytherium septentrionalis, a Fossil Edentate New to the Fauna of Texas. Journal of Mammalogy 3 (1), 1922, pp. 22-24
- ↑ George Gaylord Simpson: Holmesina septentrionalis, extinct giant armadillo of Florida. American Museum Novitates 442, 1930, pp. 1-10
- ↑ Oliver Perry Hay: Second bibliography and catalog of the fossil Vertebrata of North America. Volume II. Carnegie Institution, Washington, 1930, pp. 1–1074 (pp. 417–418)