
Replication
The replication or reduplication refers to the duplication of the DNA or RNA as the genetic information carrier of the genome or only a single chromosome or a segment of a cell or a virus . In the mitotic cell cycle, DNA synthesis takes place as an identical duplication ( reduplication ). Cell cycles of some (specialized) somatic cells in eukaryotes , however, treat parts of their genome differently: Certain DNA sequences are amplified ; other DNAs are not multiplied and remain under-replicated (in subsequent cycles) . In such cases of differentiation at the DNA level, the term replication for cellular DNA synthesis is particularly appropriate.
The molecular mechanism of replication is always semiconservative (from Latin semi 'half' and conservare 'received'). The DNA double helix is enzymatically separated into its two strands; DNA polymerases then catalyze the complementary addition to a semi-conservative (= semi-new) DNA double helix on each single strand. “Complementary” means that the isolated, conservative DNA strand clearly determines the base sequence (sequence) of its future opposite strand. According to the rules of base pairing, each base of a DNA nucleotide can only bind stably with a defined partner via hydrogen bonds ( adenine ↔ thymine ; guanine ↔ cytosine ).
In the case of RNA viruses and retroviruses , their RNA is included in the definition of replication. Because they do not have their own metabolism, all viruses use the host cells ' necessary starting materials for replication, and in some cases also their enzymes.
Semiconservative principle
Watson and Crick already recognized that the bases paired in the double strand are a prerequisite for the formation of new DNA: “It has not escaped our attention that the specific pair formation that we assume here immediately suggests a possible copying mechanism for the genetic material . “Three possibilities of replication were conceivable: dispersive or totally conservative or semi-conservative. The latter model was proven by the Meselson steel test . Accordingly, the original double strand is opened, then both single strands serve as a template in the replication (as a conservative template). The new strand is created according to the rules of Watson-Crick base pairing . The replication according to the semi-conservative principle is the generally accepted mechanism. All other principles are special cases, each of which has only been partially proven.
Since all bacteria and the cell nuclei of all eukaryotes contain double-stranded DNA (dsDNA), this replication mechanism occurs most frequently in nature. Exceptions are some mitochondria, in which a different mechanism takes place, as well as plasmid and virus genomes, in which the genetic information can be present as a single strand of DNA (ssDNA). A completely different mechanism had to be found here: the rolling circle . In retroviruses , whose genetic information is always in the form of an RNA double or single strand, replication is taken over by the host cell by transcribing the RNA into DNA using a reverse transcriptase and incorporating it into the host genome.
Summary: the replication process
In the first step, the DNA is unwound at the origin of replication by the enzyme topoisomerase . Then the double strand can be split into two single strands by the enzyme helicase (by breaking the hydrogen bonds). A replication fork is created that is held apart by SSB proteins . Primers are now attached to the single strands by the primase , as the start sequence for a DNA polymerase . The DNA polymerase reads from the 3'- to the 5'-end, synthesizing accordingly from the 5'- to the 3'-end.
The synthesis is bidirectional: Synthesis is carried out continuously on the leading strand, as it runs from 5 'to 3', whereas the following strand is discontinuous because the strands are antiparallel (in opposite directions). Okazaki fragments are therefore formed on the following strand (the strand is synthesized piece by piece by DNA polymerase III from 3 'to 5'). The primer is replaced by another DNA polymerase . The fragments are then linked again by the ligase (adhesive enzyme) to form a complete strand. The result is two identical strands of DNA; the replication took place semi-conservatively, since each double strand consists of an original and a newly synthesized single strand.
The replicon

The replicon is the elementary unit of replication. Prokaryotes and plasmids usually have only one replicon; this is sufficient to multiply the small genomes in a short time. Exceptions are some bacteria and archaea. The large genomes of the eukaryotes are divided into several or many replication segments. A replicon is usually symmetrically active, in the S phase , a time window of the cell cycle for DNA synthesis.
In the middle of the replicon is the origin , the origin of replication. As soon as the origin is activated, a replication fork (growth fork) moves in one direction and the other fork in the opposite direction. Thus, an open fork corresponds to its “twin fork”; both move away (ideally) at the same speed from the common origin. Since DNA polymerases replicate 3 '→ 5', the HO template is first available as a lead strand on one side of the original. On the other side of the origins, the other strand inevitably becomes the main strand. The replicon is fully replicated in both directions as soon as each of the two forks meets the replication fork of its neighboring replicon. A replicon end or terminus usually does not have a defined sequence, it merely requires a finite length for the replicon.
Origin
Eukaryotes have very active and less active replication start sites. There are also sleeping origins, which are intended for emergencies. The number of replication starting points corresponds to the size of the respective genome. Intestinal bacterium Escherichia coli : 1 origin. Baker's yeast Saccharomyces cerevisiae : ~ 350 origins. Human Homo sapiens : 40,000 to 80,000 origins.
Replication fork
With pulses of radioactive labeling, it was possible to visualize the DNA synthesis in the light microscope in Chinese hamster cells and HeLa cells . The chromosomal DNA consists of tandem sections (replicons) in which the (mother) DNA replicates in fork-like growth points. The progress of a fork was estimated to be 2.5 microns per minute or less. It is also important that neighboring forks become active at the same time. However, this does not exclude the possibility that the replication time of the euchromatin differs from that of the heterochromatin .
Molecular genetic studies yielded models for the submicroscopic details of the fork:
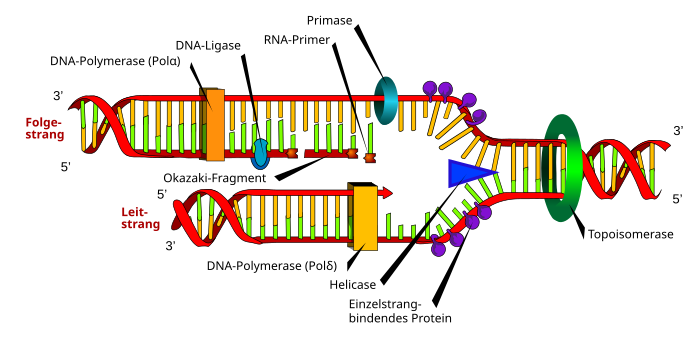

The diagram shows the enzymes or molecules involved in eukaryotes on a replication fork moving to the right from the origin. A topoisomerase works first ; it unwinds the DNA molecule from complex structures of the interphase chromatin. A helicase separates the two antipolar strands of the double helix. Protein groups stabilize the single strands so that they do not immediately pair again in a complementary manner. A DNA polymerase (Pol δ) has been continuously supplementing the leading strand 3 '→ 5' to the semi-coservative, semi-new double helix since its start at the origin .
The replication also takes place 3 '→ 5' on the next strand , but discontinuously . First, an RNA / DNA primase (Pol α) catalyzes an RNA primer which is extended to a short piece of DNA, an Okazaki fragment . A pole δ takes over the replication of the following strand to the double helix for a short distance. Then a Pol α catalyzes the next primer, the next Okazaki fragment follows, and so on. A DNA ligase with deoxynucleotides fills the gaps left by an RNase H (not shown in the scheme) when the RNA primer was degraded.
The coordinated, simultaneous progress of the following strand replication with that of the leading strand cannot be seen in the fork scheme described. So that the primase / Pol α on the next strand has the opportunity to catalyze an RNA primer and attach the first deoxynucleotides, the next strand forms a loop . In this one Pol δ molecule replaces the primase / Pol α and extends the initial RNA-DNA hybrid to an Okazaki fragment. With the help of the loop, the following strand pole δ reaches the continuously active pole δ of the leading strand in the same direction. Both polymerases combine and accomplish the replication synchronously. This model explains the importance of two other molecules. In order for the Pol δ to bind to the template, the replication factor C (RFC) is required, which also charges the PCNA (the nuclear antigen of proliferating cells). The clamp of the PCNA prevents pole δ from falling off the die.
Since the replication fork running in the opposite direction from the same origin also forms a loop for its subsequent strand, the replication eye is a tension-free, symmetrical figure.
Terminus
In eukaryotes there is no defined sequence within the chromosomes for the termination of replication; the term is only a molecular morphological description. Nevertheless: no rule without exception. There are sequence-specific barriers that stop a replication fork before the opposing fork arrives. The polar replication barriers that limit ribosomal DNA are best studied. These termini are the prerequisite for the local amplification of the rDNAs. Then the specific termination before the telomeres must be mentioned; replication stops when the fork reaches the end of the chromosome.
Replication process
- Initiation triggers replication. Here the DNA double helix is broken open at a certain point with the help of the helicase and a polymerase is attached to the broken DNA after being marked by a primase .
- Elongation , in which the actual reproduction takes place. The two strands are synthesized in a complementary manner at the same time.
- Termination completes the DNA synthesis.
- DNA repair to correct any copying errors that may have occurred.
Prokaryotic replication
initiation
In the cell, the helical DNA is not arranged in a circular or linear fashion, but is also twisted into so-called ' supercoils ': While the DNA double helix in eukaryotes wraps around basic DNA structural proteins such as histones and the entire complex of DNA Molecule and associated structural proteins are additionally compressed by folds and twisting and thus also stabilized, this is not the case with prokaryotes (as well as organelles with their own DNA such as mitochondria and chloroplasts), since they do not have histones. It has, however, histone proteins' ( English histone-like protein , HLP) found that for a similar care effect. In order to be replicated, the DNA has to be untwisted. The consequence of the unwinding of the DNA at one point is the increasing twisting of the entire DNA double strand. To counteract torsional tension during unwinding, a topoisomerase runs in front of each replication fork , which can reduce twisting (relaxation reaction). This requires the cleavage of the DNA strands. Depending on the type of enzyme (topoisomerase I or II (II is gyrase in E. coli )), single or double strand breaks are carried out in a controlled manner. After unwinding, the previously cleaved phosphoric acid ester bonds of the sugar-phosphate structure of the DNA are tied again by the enzyme.
For the initiation of replication is a special place, the origin of replication ( English Origin ) on the generally annular DNA required which determines the start point. At this point, the hydrogen bonds between the bases of the two individual strands are broken. The so-called oriC comprises 245 base pairs (bp) and contains a tandem arrangement with AT-rich sequences that have the following consensus sequence:
- 5′-GATCTNTTNTTTT-3 ′
- 3′-CTAGANAANAAAA-5 ′
There are 5 binding sites for the DnaA hexamer in E. coli. First, the initiator protein DnaA is activated by adenosine triphosphate (ATP) and bound to five 9 bp long DnaA boxes. In total, around 20 DnaA proteins are gathered in a loop around the DNA . The proteins IHF and FIS bind to specific sections of the oriC and trigger the bending of the DNA into a hairpin-like structure, which also supports the binding of DnaA. Finally, the unwinding of the double helix can start at three consecutive 13 bp long adenine- and thymine-rich sequences (also known as '13-mer sequences ').
This process is catalyzed by a helicase using ATP (in E. coli this is called DnaB and is carried to the Origin by the DnaC protein). The helicase operates from 5'-3 'direction. The separation of the double strand creates two replication forks on the origin, which diverge bidirectionally during replication. Thus, the base does not rise above hydrogen bonds mate, keep so-called single-strand binding proteins (in prokaryotes, this means "SSB protein" for English single-beach-binding protein ) apart, the individual strands.
The opening is followed by priming: an RNA polymerase , the primase (in E. coli DnaG), sets a short piece of RNA, the primer (under cellular conditions, about 10 nucleotides) on the now free single strands . This complex is called a ' primosome ' and is necessary because the main protein complex of replication, the DNA polymerase , can only begin to synthesize the second strand of DNA at a free 3'-OH group. This means that the DNA polymerase needs the primer as a 'starting aid' for replication, even if it is RNA. The RNA polymerase used for the primer only needs the single strand as a template. If the DNA polymerase has only started to synthesize the second strand (from 5 'to 3'), it can hardly be interrupted and continues to work until termination. Thus, the regulation of replication must take place in the initiation phase.
Elongation
After the initiation and the now beginning polymerization , the elongation phase is carried out. Here, the DNA polymerase III synthesizes the complementary strands to the single strands: The bases of the single strands are read one after the other and, according to the principle of complementary base pairing, incorporated one after the other in the synthesized strand. The building blocks required for DNA synthesis are available in the cell in the form of free nucleotides.
However, a problem arises here: the DNA polymerase can only synthesize in the 5 '→ 3' direction. However, since the DNA polymerase complex is synthesized on both strands at the same time, but both strands are oriented in opposite directions according to the double-helical structure , two opposing (also anti-parallel) strands result on the replication fork. After a single priming, replication can be carried out continuously on the guide strand up to the axis of symmetry of the replication fork, since this is oriented precisely with the reading direction of the polymerase complex and in the running direction of the replication fork. On the other hand, continuous replication is not possible on the following strand , since it runs in the "wrong direction". During the first run, the polymerase runs against the primer of the leading strand of the second replication fork, which runs in the other direction. After the interruption, the next strand must be primed again so that the polymerase can start again. This priming always takes place directly following the helicase. In the following irregular polymerase cycles on the following strand, the polymerase always terminates the synthesis at the last RNA primer, i.e. at the 5 'end of the preceding fragment. The resulting individual DNA fragments are also referred to as Okazaki fragments . The DNA polymerase has different domains: the actual polymerase domain for attaching nucleotides, the DNA clamp to slide on the DNA without moving and the 5'-3 ' exonuclease domain, which is used for proofreading directly after the Installation of the bases is used.
There is some well-established evidence that the area between the primase and the last Okazaki fragment is twisted into a loop (also known as the "trumpet model") so that the polymerase can work both strands in the same direction. If the synthesis of the loop is finished, it is broken up again and a new loop is formed. Here again a topoisomerase seems to be involved . Since the doubling is continuous on one strand and interrupted on the other, it is also referred to as semi-discontinuous doubling.
In order to create a continuous strand that does not contain any RNA pieces, another mechanism comes into action during replication: an RNase H removes the RNA primers and DNA polymerase I fills the resulting gap with the respective complementary DNA. DNA polymerase I can also remove RNA itself.
The DNA ligase then closes the bond from the 3 'end of the new to the 5' end of the old piece of DNA, thus establishing the phosphodiester bonds between the newly synthesized DNA strands .
Termination
In the prokaryotes with a circular DNA, termination sequences opposite the origin have been found. These are more precisely two sequences, each one for a replication fork. Normally, the termination does not have to be triggered in particular, because if two replication forks meet or the DNA ends, as in a linear form, the replication is automatically terminated. This is a control element so that the replication ends in a controlled manner at a certain point at different replication speeds of the two replication forks. The termination points are binding sites for the protein Tus ( English terminus utilizing substance ). This blocks the replicative helicase (DnaB) and thus brings replication to a standstill.
In prokaryotes, the replicated ring-shaped strands remain connected to one another for a while after replication, precisely at this terminal point, so that they can finally be separated and divided by further processes after cell division. Without this connection, there appears to be no control over the distribution. The separation of the DNA rings can take place via two mechanisms, either a type I or a type II topoisomerase being involved.
Eukaryotic replication
Replication is essentially identical in eukaryotes. However, there are some exceptions and special cases. It must be taken into account that the DNA is more “packaged” (e.g. in the case of heterochromatin ), the DNA-binding proteins (the histone and non-histone proteins ) have a stronger influence and the DNA is in a linear form. In addition, the proteins involved are usually those with the same functionality but different structure.
One of the main differences lies in the initiation : there are several origins among eukaryotes. The advantage of this is easy to see because, on the one hand, the replication is slower due to the more DNA-binding proteins present, and on the other hand, the eukaryotic polymerase (different polymerases, which are designated with Greek letters and which are involved in the synthesis of leading and subsequent strands - Polα respectively Polδ - differentiate), which has a more complex repair mechanism than in prokaryotes ( English proofreading ), progresses more slowly. In eukaryotes, polymerase creates around 50 to 100 nucleotides per second, while in prokaryotes more than 1000 nucleotides per second can be added. Furthermore, the eukaryotic DNA is usually much larger than that of the prokaryotes (several million base pairs in prokaryotes compared to a few billion in eukaryotes). Several origins and thus several replication units shorten the time that is required to replicate the entire genome, even if the speed of prokaryotes is not reached (the replication time of prokaryotes is in the range of a few minutes, for eukaryotes it is several hours).
The origins of the eukaryotes do not have a special sequence, rather it is a so-called consensus sequence , i.e. a similarity sequence . These are also called ARS elements . Other findings assume that large areas of DNA, so-called replication centers, can serve as possible starting points for replication. The, however, as Origin recognized sites are inactivated by an Origin recognition complex ( English origin recognition complex , ORC), a Cdc6 protein and so-called MCM proteins ( English minichromosome maintenance protein ), which serve as helicases marked. These proteins, which are later removed, form the vanguard of replication. The ORC binds to the origin, the ORC also recruits other factors (Cdc6, Cdt1 and 'Helicase Loading Proteins'), whereupon MCM helicases bind, which melt the DNA. Replication is initiated only once during the S phase (despite the 10,000 origins on the eukaryotic genome). Cyclins give the signal for replication in the cell cycle.
The further course of initiation and elongation is functionally identical to that of the prokaryotes.
Terminal sequences have not yet been discovered in eukaryotes. They don't seem to matter either, as the replication apparatus automatically stops once the end of the DNA is reached. In contrast to the ring-shaped DNA structure of the prokaryotes, a problem arises here: The DNA polymerase synthesizes on the mother strand from 3 'to 5'. (So the daughter strand has the orientation 5 '→ 3') The polymerase needs a primase as a starting aid to be able to duplicate DNA. Primase is an enzyme that replicates a short DNA start sequence as RNA. This starting piece presents the DNA polymerase with a nucleotide with a free 3'-OH end at which it can further synthesize DNA nucleotides. After successful synthesis, the RNA primers are destroyed by enzymes (flap endonucleases) and thus leave gaps. However , these gaps cannot be closed at the telomeres , the ends of the chromosomes, as there is no preceding 3 'end. The Okazaki fragments are created in a very similar way during the synthesis of the delay strand. Here the polymerase has to work in the wrong direction, which is why many primases have to be used. In eukaryotes, in contrast to prokaryotes, the primase is not present as a separate enzyme, but bound to the DNA polymerase alpha (Polα). The resulting gaps are filled by Polδ and connected with DNA ligases . This is possible because there is always a preceding nucleotide with a 3 'end. Since there are no such nucleotides on the telomeres, complete synthesis of the ends is not possible. With every chromosome duplication, the telomeres at the 5 'end of both daughter strands shorten. Since telomeres are made up of a tandem-like repetitive sequence , i.e. a sequence that is repeated one after the other and does not contain any structural genes, a loss up to a certain length is not a major disadvantage. It is assumed, however, that the more the number of replications increases, the more unstable DNA becomes, since the stabilizing effect of the telomeres becomes weaker and weaker. This could possibly be a genetic indicator of aging .
Apart from protozoa such as the eyelash animal Tetrahymena , an enzyme called telomerase has been discovered in multicellular cells in the germ line cells and stem cells as well as in the bone marrow ( blood formation ) and also in some tumor cells , which compensates for this loss. This is a reverse transcriptase (RNA-dependent DNA polymerase), because it contains the repetitive sequence as a template in RNA form. It extends the leading strand by a few sequences so that the DNA polymerase can synthesize the next strand after priming.
During the S phase of the cell cycle, the protein cohesin binds the two sister chromatids to one another along their entire length. During the anaphase , the enzyme separase dissolves the cohesin again so that the sister chromatids can be drawn from the spindle fibers to the cell poles.
Replication is triggered in a section of the cell cycle : in eukaryotes in the S phase (DNA synthesis phase ), which itself belongs to the interphase.
Reduplication in germline and stem cells
In order to preserve the genetic blueprint of a biological species and the program for its timely realization, the cells of the germ line fulfill one necessity: They have to exactly duplicate their genetic information before each nucleus and cell division. The chromosomes realize this requirement, which requires a special solution at their ends, the telomeres . (Pluripotent) stem cells also obey the strict rule for the mitotic cell cycle . Cell division is strongly linked to identical, complete, semiconservative replication .
Replication in somatic cell nuclei
- Endoreplication : In the soma of many organisms there are several successive cell cycles with genetic multiplication, after which the enlarged cell nuclei, and consequently the cells concerned, do not divide. This type of repeated DNA synthesis is called endoreplication . This may often be a complete reduplication; but often endoreplication is a selective process , namely:
- During amplification , certain DNA sequences are over-replicated compared to the rest of the genome. The chorionic genes in Drosophila melanogaster amplify sixty-fold in the follicle cells before their transcription begins. Such over-replication guarantees a large amount of mRNA for the envelope of the fly eggs.
- Under- replication, on the other hand, excludes certain sequences from the doubling rounds. The genetic result can be compared with that of the elimination. In polytene chromosomes of D. melanogaster , individual replication forks get stuck in 20% of their bands: replication is not terminated in places. However, this is a harmless type of genome instability , since it occurs in somatic cells, namely in the salivary glands.
- Diminution, elimination: Various eukaryotes forego a considerable part of their DNA in a regulated manner after a certain embryonic development point. Cases in which chromosomes are wholly or partly removed from somatic cell nuclei look particularly dramatic under the light microscope. The technical terms for this are chromatin diminution or chromosome elimination .
Structural Mechanisms
Replication can take place with various molecular mechanisms, which depend on the primary structure of the nucleic acid. In addition to the symmetrical process of bidirectional replication, which has been described so far, there are asymmetrical processes, namely: telomere reproduction, D-loop process and rolling circle principle.
Bidirectional replication
The replication of the DNA in linear chromosomes of the nuclear genome of a eukaryotic cell and of the bacterial genome takes place symmetrically on both sides. Replication is based on many starting sequences ("origins") that are distributed over the DNA double helix (of a chromosome). The number of origins corresponds to the (possible) number of replication units (“replicons”). The bidirectional replication runs in the opposite direction from each origin, in both directions, and at the same time on both single strands of the DNA double helix. The bidirectional principle occurs most frequently in nature.
Telomere reproduction
The identical replication of the telomeres is a problem with linear DNA molecules or their chromosomes. The two ends of a double helix do not allow bidirectional replication because there is ultimately no starting sequence for an RNA primer on the “subsequent strand”. This is why the following strand at the end of the chromosome remains a primer length (20 to 200 nucleotides) shorter than the “leading strand”. Since normal DNA replication is not possible, telomerase starts at the lead strand . This (hybrid) enzyme consists of a protein part and the RNA component 3' - C AACCCC AA-5 '(in Tetrahymena ). The nine RNA nucleotides contain the template (bold), which accordingly the single-stranded telomere segment 5'-TTGGGG-3 'is repeatedly strung together and the (former) leading strand is extended in tandem.
- Telomerase: The enzyme acts on the 5'-3 'DNA template as reverse transcriptase , in three steps: 1. Docking with the RNA sequence at the 3' end of the chromosomal DNA leading strand; 2. Extending the 3 'end; 3. Advance one telomeric motif to the 3 'end just formed. It works like the construction of a bridge that is driven forward with a self-supporting structure. Finally, the single-stranded telomere DNA flips over a little and forms extraordinary GG base pairs with itself. In the resulting loop, the single-stranded DNA is completed by the (normal) DNA polymerase to form a double helix.
Cells with active telomerase are able to reproduce and preserve the telomeres before each division. This is particularly necessary in the germ line and in stem cells. Aging somatic cells no longer meet this requirement when their telomeres “wear out”.
- Telomer loss: As far as the facts can be measured, the cell nuclei contain shorter telomeres after each division. The loss of repetitive telomer sequences is part of the cellular aging process and is particularly noticeable in connection with many diseases. If the telomer sequences are no longer fully reproduced, the chromosome ends will eventually no longer be adequately protected. This leads to genomic instability.
- Telomere transposition: The discovery that a model organism lacks telomerase was a surprise. The fruit flies of the species Drosophila complete the chromosomal telomeres by transposition . While the Drosophila species have lost the telomerase gene, it is still present in other organisms with telomere transposition; these include the silkworm Bombyx mori and the red-brown rice flour beetle Tribolium castaneum .
D-loop process
Some chloroplasts and mitochondria have circular DNA. Replication begins on one strand in the large, non-coding region. The polymerase complex runs in one direction only, producing a short, third strand known as 7S DNA. This three-strand structure can be seen in the electron microscope as a D-loop . (D-Loop from displacement loop : detachment or displacement loop .) If the already replicated strand has displaced more than two thirds of the original DNA, it becomes detached (from the template). The loosened strand is replicated independently (to the dsDNA).
Rolling circle replication

Viruses and plasmids show a special principle of replication. If their nucleic acid is present as a single strand, it is complemented to form a double strand. If the genetic material is (already) a double-stranded nucleic acid, an endonuclease breaks one strand by breaking the connection between two neighboring bases. A polymerase complex that only works in one direction comes into play at this open point . The 3'-OH end of the cut strand serves as a starting point for a so-called primer from which the open strand is extended. The unbroken ring serves as a complementary template for one-sided polymerization . The replication unit travels around the inner strand around like a rolling circle ( Rolling circle ).
The inner strand can repeatedly serve as a template, so that several duplicates arise one after the other, sometimes as concatamers . These are broken down after the first replication step. The new products either remain single-stranded or serve as a template for a second step from which the replicated double-stranded nucleic acids arise.
The rolling circle principle seems to exist in nature in different forms. The process described represents the most widespread hypothesis. - Explanation of viruses: These can be divided into six classes according to the type of their nucleic acids. 1. Viruses with double-stranded DNA; 2. with single-stranded DNA; 3. with double-stranded RNA; 4. with positive RNA single strand; 5. with negative RNA single strand; 6. Retroviruses replicate their RNA to DNA, from which new virion RNA is eventually multiplied.
The rolling circle model occurs, for example, when two bacteria are conjugated . A bacterium passes on the single strand of a plasmid to another bacterium, while it retains its own, circular DNA of the plasmid. As yet little researched circular DNAs have been observed extrachromosomally in human cancer cells.
See also
literature
- Rolf Knippers, Alfred Nordheim (Ed.): Molecular Genetics . 10., completely revised. and exp. Edition. Thieme, Stuttgart 2015, ISBN 978-3-13-477010-0 .
- Inge Kronberg: Genetics, 9 - DNA: Carrier of genetic information. Pages 147-158. In: Jürgen Markl (ed.): Markl Biologie Oberstufe. Ernst Klett, Stuttgart, Leipzig 2010. ISBN 978-3-12-150010-9 .
- Wilhelm Seyffert: Textbook of Genetics. Gustav Fischer, Stuttgart etc. 1998. ISBN 3-437-25610-6 . → Chapter 4: Transmission of genetic information. Pages 59-80.
- Melvin L DePamphilis (ed.): DNA replication in eukaryotic cells. Cold Spring Harbor USA, CSH Laboratory 1996. ISBN 0-87969-459-9 .
Web links
- David Hull and John S. Wilkins: Entry in Edward N. Zalta (Eds.): Stanford Encyclopedia of Philosophy .
- Maxanim animated representation of DNA replication
- Replication of DNA with pictures and description
- Animated representation of the DNA replication with explanations
- "For the first time, researchers are recording how DNA doubles - and it is different than expected," Galileo.tv, June 2016
Individual evidence
- ↑ a b R. Chaudhry, SS Bhimji: Biochemistry, DNA Replication. StatPearls Publishing; 2018. PMID 29489296 .
- ↑ Craig E. Cameron: Viral Genome Replication. Springer Science & Business Media, 2009, ISBN 978-0-387-89456-0 , p. 201.
- ↑ Tai-An Cha, Bruce M. Alberts : In vitro studies of the T4 bacteriophage DNA replication system. In: Thomas Kelly , Bruce Stillman (eds.): Cancer Cells. 6: Eukaryotic DNA replication. CSH Laboratory, Cold Spring Harbor 1988: 1-10. ISBN 0-87969-308-8 .
- ↑ James D Watson : The Double Helix: A Personal Account of the Discovery of DNA Structure. Rowohlt, Reinbek near Hamburg 1969; Page 170. ISBN 3-499-16803-0 .
- ↑ Gunther S Stent (ed.): James D Watson: The double helix. A personal account of the discovery of the structure of DNA. Norton & Company, New York, London. ISBN 0-393-95075-1 . Page 129: "It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material."
- ^ Matthew Meselson , Franklin W Stahl : The replication of DNA in Escherichia coli. In: Proceedings of the National Academy of Sciences . Volume 44, Number 7, July 1958, pp. 671-682, doi : 10.1073 / pnas.44.7.671 , PMID 16590258 , PMC 528642 (free full text).
- ↑ József Szeberényi: The meselson steel experiment. In: Biochemistry and Molecular Biology Education. 40, 2012, p. 209, doi : 10.1002 / bmb.20602 . ← Questions about understanding the experiment.
- ↑ François Jacob , Sydney Brenner : Sur la régulation de la synthèse du DNA chez les bacteries: l'hypothèse du replicon. In: CR Acad Sci Paris 256, 1963: 298-300.
- ↑ Marcelina W Musiałek, Dorota Rybaczek: Behavior of replication origins in Eukaryota - spatio-temporal dynamics of licensing and firing. In: Cell cycle. Volume 14, number 14, 2015, pp. 2251-2264, doi : 10.1080 / 15384101.2015.1056421 , PMID 26030591 , PMC 4614997 (free full text) (review).
- ↑ Rachel L Creager, Yulong Li, David M MacAlpine: Snapshot: Origin of DNA-replication. In: Cell 161, April 2015.
- ^ Marie-Noëlle Prioleau, David M MacAlpine: DNA replication origins - where do we begin? In: Genes & development. Volume 30, number 15, 08 2016, pp. 1683–1697, doi : 10.1101 / gad.285114.116 , PMID 27542827 , PMC 5002974 (free full text) (review).
- ↑ Joel A Huberman, Arthur D Riggs: On the mechanism of DNA replication in mammalian chromosomes. In: J Mol Biol 32, 2, 1968: 327-334. Abstract.
- ↑ Shou Waga, Bruce Stillman : Anatomy of a DNA replication fork revealed by reconstitution of SV40 DNA replication in vitro. In: Nature 369, 6477, 1994: 207-212. DOI: 10.1038 / 369207a0. → Loop model: p. 211, Fig. 6.
- ↑ Shou Waga, Bruce Stillman: The DNA replication fork in eukaryotic cells. In: Annu Rev Biochem 67, 1998: 721-751. PDF.
- ↑ Agnieszka Gambus: Termination of eukaryotic replication forks. In: Adv Exp Med Biol 1042, 2017: 163-187. doi: 10.1007 / 978-981-10-6955-0_8.
- ↑ James M Dewar, Johannes C Walter: Mechanisms of DNA replication termination. In: Nature reviews. Molecular cell biology. Volume 18, number 8, 08 2017, pp. 507-516, doi : 10.1038 / nrm.2017.42 , PMID 28537574 , PMC 6386472 (free full text) (review).
- ↑ A Bębenek, I Ziuzia-Graczyk: Fidelity of DNA replication - a matter of proofreading. In: Current Genetics. [Electronic publication before going to press] March 2018, doi: 10.1007 / s00294-018-0820-1 , PMID 29500597 .
- Jump up ↑ Neil A Campbell, Jane B Reece, Lisa A Urry, Michael L Cain, Steven A Wasserman, Petzer V Minorsky, Robert B Jackson: Biology. 8th edition. Benjamin Cummings, Pearson Switzerland, Cham 2008. ISBN 978-0-321-54325-7 .
- ↑ John M. Archibald: The Puzzle of Plastid Evolution . In: Current Biology . 19, No. 2, 2009, pp. R81-8. doi : 10.1016 / j.cub.2008.11.067 . PMID 19174147 .
- ↑ T. Kobayashi, M. Takahara, SY Miyagishima, H. Kuroiwa, N. Sasaki, N. Ohta, M. Matsuzaki, T. Kuroiwa: Detection and Localization of a Chloroplast-Encoded HU-Like Protein That Organizes Chloroplast Nucleoids . In: The Plant Cell Online . 14, No. 7, 2002, pp. 1579-1589. doi : 10.1105 / tpc.002717 . PMID 12119376 . PMC 150708 (free full text).
- ↑ Hans-Georg Koch, Jan Brix, Peter C. Heinrich: Replication - The doubling of DNA . In: Löffler / Petrides Biochemistry and Pathobiochemistry (= Springer textbook ). Springer, Berlin, Heidelberg 2014, ISBN 978-3-642-17972-3 , pp. 545-558 , doi : 10.1007 / 978-3-642-17972-3_44 .
- ^ A b Replication and repair mechanisms of DNA - knowledge for medical professionals. Retrieved February 18, 2020 .
- ↑ BA Berghuis, VS Raducanu, MM Elshenawy, (…), NH Dekker: What is all this fuss about Tus? Comparison of recent findings from biophysical and biochemical experiments. In: Crit Rev Biochem Mol Biol 53, 2018: 49-63. doi: 10.1080 / 10409238.2017.1394264.
- ^ Carol W. Greider , Elizabeth H. Blackburn : Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. In: Cell . tape 43 , 1985, pp. 405-413 , doi : 10.1016 / 0092-8674 (85) 90170-9 , PMID 3907856 .
- ^ Lynne Suzanne Cox: Molecular Themes in DNA Replication. Royal Society of Chemistry, 2009, ISBN 978-0-85404-164-0 , p. 156.
- ↑ Teresa Rivera, Candy Haggblom, Sandro Cosconati, Jan Karl Eder: A balance between elongation and trimming Regulates telomere stability in stem Cellls. In: Nature structural & molecular biology. Volume 24, number 1, 01 2017, pp. 30-39, doi : 10.1038 / nsmb.3335 , PMID 27918544 , PMC 5215970 (free full text).
- ↑ Julia Su Zhou, Eros Lazzerini Denchi: How stem cells keep telomeres in check. In: differentiation; research in biological diversity. Volume 100, 2018 Mar - Apr, pp. 21-25, doi : 10.1016 / j.diff.2018.01.004 , PMID 29413749 , PMC 5889314 (free full text) (review).
- ↑ Hewson Swift : The constancy of desoxyribose nucleic acid in plant nuclei. In: Proc Nat Acad Sci USA 36, 1950: 643-654.
- ↑ ER Gaginskaya, VL Kasyanov, GL Kogan: Amplification of ribosomal genes and formation of extrachromosomal nucleoli in oocytes of starfish Henricia hayashi (Asteroidea: Echinasteridae). In: Cell Differ 23, 1988: 53-60.
- ↑ Ulrich Scheer, Michael F Trendelenburg, G Krohne, Werner W Franke : Lengths and patterns of transcriptional units in the amplified nucleoli of oocytes of Xenopus laevis. In: Chromosoma 60, 2, 1977: 147-167.
- ↑ Allan C Spradling : The organizattion and amplification of two chromosomal domains containing Drosophila chorion genes. In: Cell 27, 1981: 193-201.
- ↑ Terry L Orr-Weaver, Allan C Spradling: Drosophila chorion gene amplification requires an upstream region regulating s18 transcription. In: Molecular and Cellular Biology . Volume 6, Number 12, December 1986, pp. 4624-4633, doi : 10.1128 / mcb.6.12.4624 , PMID 3099171 , PMC 367247 (free full text).
- ↑ EN Andreyeva, TD Kolesnikova, ES Belyaeva, RL Glaser, Igor F Zhimulev: Local DNA underreplication correlates with accumulation of phosphorylated H2Av in the Drosophila melanogaster polytene chromosomes. In: Chromosome Res 16, 2008: 851-862.
- ↑ Helmut Zacharias: Allocyclic behavior and underreplication of the nucleolus chromosome in Pseudodiamesa (Chironomidae). In: Chromosoma 89, 1984: 263-273.
- ^ Emil Heitz : About α- and β-heterochromatin as well as constancy and structure of the chromomers in Drosophila. In: Biol Zentralblatt 54, 1934: 588-609. → Page 596: Describes underreplication of heterochromatin for the first time in Drosophila virilis .
- ↑ Allan C Spradling: Polytene chromosome structure and somatic genome instability. In: Cold Spring Harb Symp Quant Biol 82, 2017: 293-304. PDF.
- ^ Theodor Boveri : The development of Ascaris megalocephala with special consideration for the core conditions. In: Festschrift for the seventieth birthday of Carl von Kupffer . Fischer, Jena 1899: 383-429.
- ↑ Theodor Boveri: The potencies of the Ascarisblastomere with modified furrowing. At the same time a contribution to the question of qualitatively unequal chromosome division. In: Festschrift, Richard Hertwig's 60th birthday . Vol III. Fischer, Jena 1910: 133-214.
- ^ Sigrid Beermann: The diminution of heterochromatic chromosomal segments in Cyclops (Crustacea, Copepoda). In: Chromosoma 60, 1977: 297-344.
- ^ RA Finch: Tissue-specific elimination of alternative whole parental genomes in one barley hybrid. In: Chromosoma 88, 1983: 386-393.
- ^ Walter Nagl: Heterochromatin elimination in the orchid Dendrobium. In: Protoplasma 118, 1983: 234-237.
- ^ Dieter Ammermann: Germ line specific DNA and chromosomes of the ciliate Stylonychia lemnae. In: Chromosoma 95, 1987: 37-43.
- ↑ Jianbin Wang, Shenghan Gao, Yulia Mostovoy, Yuanyuan Kang, Maxim Zagoskin, Yongqiao Sun, […], Richard E Davis: Comparative genome analysis of programmed DNA elimination in nematodes. In: Genome research. Volume 27, number 12, 12 2017, pp. 2001-2014, doi : 10.1101 / gr.225730.117 , PMID 29118011 , PMC 5741062 (free full text).
- ↑ Elizabeth H Blackburn: Telomerases. In: Annu Rev Biochem 1992.61, 1992: 113-129. PDF.
- ↑ Carol W Greider, Elizabeth H Blackburn: A telomere sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. In: Nature 337, 6205, 1989: 331-337. → Page 335: "The elongation translocation mechanism."
- ↑ Helmut Zacharias, Inge Kronberg: Telomeres: All's well that ends well. In: Biologie in our Time 6, 2009: 366–367.
- ↑ David Van Ly, Ronnie Ren Jie Low, Sonja Frölich, Tara K Bartolec, Georgia R Kafer, Hilda A Pickett, Katharina Gaus, Anthony J Cesare: Telomere loop dynamics in chromosome end protection. In: Mol Cell 71, 4, 2018: 510-525.e6. PDF.
- ↑ Elizabeth H Blackburn, Carol W Greider, Jack W Szostak : Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. In: Nature Med 12, 2006: 1133-1138. DOI: 10.1038 / nm1006-1133.
- ↑ Kara J Turner, Vimal Vasu, Darren K Griffin: Telomere biology and human phenotype. In: Cells. Volume 8, number 1, 01 2019, p., Doi : 10.3390 / cells8010073 , PMID 30669451 , PMC 6356320 (free full text) (review).
- ↑ Elena Casacuberta, Mary-Lou Pardue: HeT-A elements in Drosophila virilis: Retrotransposon telomeres are conserved across the Drosophila genus. In: Proceedings of the National Academy of Sciences . Volume 100, number 24, November 2003, pp. 14091-14096, doi : 10.1073 / pnas.1936193100 , PMID 14614149 , PMC 283551 (free full text).
- ↑ Elena Casacuberta: Drosophila: Retrotransposons making up telomeres. In: Viruses. Volume 9, number 7, 2017, doi : 10.3390 / v9070192 , PMID 28753967 , PMC 5537684 (free full text) (review). → Page 10: Compares similarities between telomere retrotransposons and the telomerase mechanism.
- ↑ Thomas J Nicholls, Michal Minczuk: In D-loop: 40 years of mitochondrial 7S DNA. In: Exp Gerontol 56, 2014: 175-181. → The function of the 7S-DNA is still up for discussion here.
- ↑ Josef Köck, Christine Rösler, Jing-Jing Zhang, Hubert E Blum, Michael Nassal, Christian Thoma: Generation of covalently closed circular DNA of hepatitis B viruses via intracellular recycling is regulated in a virus specific manner. In: PLoS pathogens. Volume 6, number 9, September 2010, p. E1001082, doi : 10.1371 / journal.ppat.1001082 , PMID 20824087 , PMC 2932716 (free full text).
- ↑ JA Ruiz-Masó, C Machón, L Bordanaba-Ruiseco, M Espinosa, M Coll, G Del Solar: Plasmid rolling-circle replication. In: Microbiol Spectr 3, 1, 2015: PLAS-0035-2014. PDF.
- ↑ T Santos, P Pereira, JA Queiroz, C Cruz, F Sousa: Plasmid production and purification: An integrated experiment-based biochemistry and biotechnology laboratory course. In: Biochem Mol Biol Educ 2019 Nov; 47, 6, 2019: 638-643.
- ^ Salvador Edward Luria , James E. Darnell Jr., David Baltimore , Allan Campbell: General Virology. 3rd edition. John Wiley & Sons, New York etc. 1978. ISBN 0-471-55640-8 . → Pages 306f: Viral genetic systems: Classification of animal viruses.
- ↑ Eliszabeth Pennisi: Circular DNA throws biologists for a loop. In: Science 356, 6342, 2017: 996.