
Tardigrade
| Tardigrade | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

The tardigrade Milgiene tardigradum |
||||||||||||
| Systematics | ||||||||||||
|
||||||||||||
| Scientific name | ||||||||||||
| Tardigrada | ||||||||||||
| Spallanzani , 1777 | ||||||||||||
| Classes | ||||||||||||
Tardigrades (Tardigrada) - also called water bears - form an animal strain within the molting animals ( Ecdysozoa). The eight-legged animals, usually less than a millimeter in size, are reminiscent of bears due to their appearance and their clumsy-looking way of locomotion , which led to their name in German-speaking countries . Its scientific name (a combination of the Latin tardus 'slow' and gradus 'step' ) goes back to the slow movement.
They live worldwide in the sea, fresh water or in humid habitats on land; they are particularly common there in moss pillows . One characteristic of animals is cryptobiosis , a death-like state in which they can survive extreme environmental conditions. Tardigrades can feed on the contents of plant cells as well as predatory on small animals such as roundworms (Nematoda) or rotifers (Rotifera), which they prick and suck out. Tardigrades usually reproduce sexually . Some species also reproduce parthenogenetically , that is, without the participation of males; the eggs of the females in this case develop without fertilization.
The next recent relatives of tardigrades are probably limbs (Arthropoda) and velvet worms (Onychophora) with which they the taxon Panarthropoda form.
construction

The size of tardigrade is between 50 micrometers for some juveniles and 1.5 millimeters for a species of the genus Macrobiotus , but is usually around 100 to 500 micrometers, so that the animals are counted as meiofauna . They have a relatively plump, cylindrically shaped body that is flattened on the abdomen and, seen on the surface, appears to consist of four body segments, each with a pair of legs and a head segment. This external impression, caused by transverse folds in the outermost layer of skin, is misleading, however: the head, which is only slightly separated and differentiated from the rest of the body, probably consists not of one, but of three fused segments; the body, on the other hand, could not consist of four, but possibly of be composed of five parts.
Marine species are mostly colorless or inconspicuously white to gray in color, while land and freshwater species often have colors such as red, yellow, green, pink, purple or black. The coloring is caused either by pigments in the outer skin, the cuticle , or by the colored contents of the body cavity, the hemocoeloma , or the intestine. Dyes often also migrate from the intestine into the haemocoeloma and from there are deposited in the cuticle.
Characteristically, many of the tardigrade tissues are ugly , that is, the number of cells they consist of is genetically determined. Growth in size cannot therefore take place through an increase in the number of cells, but only through growth of the individual cells themselves. If one of the two daughter cells is killed (by experimental intervention) after the first cell division, an anatomically complete and viable animal develops that then contains only half as many cells as normal.
extremities

On four trunk segments, on the left and right edge of the abdomen (ventrolaterally), a "stump leg" arises, a jointless protuberance of the trunk, as is also found in the related group of stumpefoots . Occasionally, a distinction is made between individual regions of the leg, which are then referred to as the coxa ("hip"), femur ("thigh"), tibia ("shin") or tarsus ("foot"). Since the legs are only slightly differentiated, the anatomical basis of these terms is questionable; In any case, however, they are to be understood purely descriptively, i.e. do not indicate an evolutionary relationship with the corresponding leg components of the arthropods.
Many marine species are able to telescope their legs into one another; The short muscles running within the leg of all tardigrade help them in this. At the end of the leg there are usually four to eight, more rarely up to thirteen retractable claws, which sometimes sit on thin leg growths, the "toes", and are secreted by special glands, the "claw glands" . In some species the claws are secondary to adhesive disks; there are no claws in the genus Apodibius . Often the claws of the fourth pair of legs point in the opposite direction as those of the first three pairs.
skin

The body is delimited to the outside by a non-cellular outer layer, the cuticle , and an underlying inner layer consisting of individual cells, the epidermis . Flagellated cells do not appear in the tardigrade skin.
The cuticle is 0.5 micrometers thick and consists of three layers, the outer epicuticle with a thickness of about 0.1 micrometers, an underlying intracuticle with a thickness of about 0.2 micrometers and a final procuticle that is about 0 .25 microns. It is composed of chitin , various glycoproteins , polysaccharides , mucopolysaccharides , lipids and lipoproteins and also contains pigments in many terrestrial and freshwater species. Often it is ornamented with bumps, thorns or dimples and in some species it is hardened on the back (dorsal), more rarely on the side (lateral), very rarely also on the abdomen (ventral) to form thick armor plates, the sclerites ; these are then articulated as with the arthropods. The cuticle, along with the leg claws and the lining of the foregut and hindgut, including the mouthparts contained in the former, the stilettos, is regularly skinned .
The epidermis consists of a Eutelian, i.e., within a species, always consisting of the same number of cells, which is secreted by the cuticle above. It is separated from the body cavity only by a thin extracellular separating layer, the so-called basal lamina.
Hemocoeloma
The body cavity is not a true coelom , that is, it is not lined by a tissue that can be traced back to the embryonic mesoderm . All organs are separated from it only by the thin basal lamina. The body cavity is therefore referred to as a pseudo- or, since it is filled with “blood”, as a hemocoeloma . A real coelom is only found around the single gonad (gonad).
The blood is colorless and does not serve to transport oxygen. It circulates through body movements; however, there is no real blood circulation, just like separate blood vessels or a heart. For this, it is filled with up to 200 cells, the hemocytes, either floating freely or anchored to the basal lamina. Their first task appears to be the storage of nutrients; they may also play a role in combating bacteria.
Musculature
Unlike the closely related columbus, muscles in the tardigrade never appear in thick muscle layers. Instead, the muscles of the trunk and limbs consist of isolated, thin longitudinal muscles attached to the cuticle, which consist of only a few, sometimes even a single muscle cell. They either work against the haemocoel, which acts like a fluid-filled balloon as a hydroskeleton, or antagonistically, like the flexor and extensor muscles in humans, against each other. Ring muscles are not found in tardigrade. The muscle cells are mostly smooth, rarely striated, the latter presumably being the original state; According to a widespread hypothesis, smooth muscles developed in many species only as a secondary adaptation to rural life.
breathing
Muscles need oxygen for their work , the average consumption of which by tardigrade is estimated to be between 0.05 and 0.1 cubic millimeters per hour and milligram of body weight. Due to the very favorable ratio of body surface and volume, however, no separate respiratory organs are necessary and accordingly are not available. The entire gas exchange can take place through simple diffusion through the skin, which, however, must be covered by a thin film of water.
Digestive organs
The digestive tract consists of a long tube that runs from the front to the back of the body and can be divided into the oral cavity, throat (pharynx), esophagus, midgut and rectum. The first three sections are grouped together as the foregut (esophagus) because the lining of the cuticle, together with the cuticular lining of the hindgut, is shed with each molt.
In carnivorous or omnivorous species (carnivores and omnivores), the mouth opening is usually at the end of the front end of the body (terminal), in species that feed on plants or organic waste (herbivores and detritivores), on the other hand, it is often a little behind the front end Ventral side (ventral-subterminal). It is often surrounded by a ring of 10 to 30 hard, cuboidal projections, the so-called lamellae, and sometimes sits on a mouth cone that can be telescoped out.
A salivary gland opens into the adjoining tubular oral cavity on the left and right, which not only secretes digestive secretions, but also synthesizes the stilettos characteristic of tardigrade. These are hard, mostly straight, but occasionally also curved needles, which are usually located in the interior (lumen) of the respective salivary gland, but can protrude into the oral cavity with their sharp tip. Pro- and retractor muscles, which attach to their thickened rear ends, can be stretched or drawn in. Transverse stiletto holders connect them to the oral cavity, while their front end rests in their own stiletto sheaths when retracted. In carnivorous species they are usually somewhat more pronounced than in herbivorous forms.
The pharynx consists of striated epithelial muscle cells that run radially outwards and are separated from the body cavity by a basal lamina . The interior of the throat, the lumen, on the other hand, is lined with cuticles and, thanks to its Y-shaped, triadidic cross-section, is ideally suited for the efficient generation of negative pressure, which is used to suck liquid food into the intestine. This pump principle has also developed independently in other, not closely related animals such as rotifers .
A short esophagus consisting of cubic cells, which presumably secretes mucus, connects to the midgut, which consists of a single layer of epithelial tissue that sometimes has lateral protuberances (microvilli), which are believed to serve to increase the surface area. When entering the intestine, the food is often enveloped by a peritrophic membrane , the function of which is to protect the sensitive intestinal tissue from damage by foreign bodies. The midgut secretes hydrolytic enzymes , absorbs nutrients from food, stores them in the form of fats or polysaccharides and probably also has a function in the elimination of waste materials. In many species there is an extensive bacterial flora that presumably lives commensally , i.e. without influencing its host, but in some cases also serves as a source of food.
The food residues are collected in a short rectum lined with a cuticle, possibly modified again, and then released to the outside world through the anus on the abdomen, slightly in front of the last pair of legs on the midline (medial-ventral) . In many species not only the Malpighian glands, described in more detail below, open into the rectum, but also the fallopian tubes of the females; in this case the last section of the intestine is called the cloaca.
Excretory organs
Indigestible residues, metabolic waste products or pollutants can be released to the outside world in various ways. On the one hand, many pollutants are built into the overlying cuticle by the epidermis and then shed together with it during the next moult . In addition, the salivary glands apparently take on an excretory function during the moult. The intestine not only absorbs nutrients, but also releases harmful substances from the body cavity to the intestinal lumen, the intestinal cavity.
Many species that are assigned to the class Eutardigrada also have three, very rarely four, specialized organs, which are known as Malpighian glands . Two of them are on the side (lateral), one is on the back (dorsal) of the intestine. They open at the junction of the middle and rectum and are most likely used to eliminate nitrogenous waste. Although they are structurally similar to the glands of the same name in insects, they are probably not homologous organs, which means that in evolutionary terms they do not go back to a common predecessor structure. Since they are not present in the originally considered marine species of the second great class, the heterotardigrada , they are considered an evolutionary adaptation to life in freshwater and on land.
Some rural forms of the heterotardigrada have, instead of Malpighian glands, special organs located on the abdomen, which open at the attachment of the second and third pair of legs and presumably have an excretory function. Real nephridia , highly developed excretory organs, on the other hand, do not occur in tardigrade.
Nervous system
The tardigrade's nervous system consists of a ring of nerves in the head running around the foregut and two pairs of nerve cords running backwards on the abdomen, which form a so-called rope ladder nervous system through cross-connections in each trunk segment .
The front nerve ring consisting of a lying above the mouth supraesophageal ganglion and one located below the same subesophageal , two clusters of nerve cells, which passes through the side of the digestive tract nerve tapes are connected to each other to form a ring, and as a primitive " brain can be considered". The upper pharyngeal ganglion consists of two pairs of brain lobes located on the rear and facing the rear end (caudal), an inner and an outer pair, as well as a further pair located on the side of the stilettos. The outer rear pair, if present, innervates the eyes and also supplies other sensory organs located on the head, the cirri and clavae, with nerves. It is also in an unusual way connected to the ganglia of the first trunk segment. As with the related arthropods, however, these are primarily connected to the subterranean ganglion by broad nerve ligaments.
The tardigrade brain arises from the fusion of several originally independent ganglia, which can be interpreted as an indication of the origin of the head from the fusion of at least three segments. The brain of the closely related arthropods is also composed of several ganglia, which there form the three brain regions proto-, deuto- and tritocerebrum. This is why it is sometimes assumed that the corresponding structures are homologies , i.e. tissues that go back to common ancestors of tardigrade and arthropod. More recent electron microscopic studies contradict this view and instead come to the conclusion that the entire brain of the tardigrade corresponds in evolutionary terms to the protocerebrum of the arthropod.
Two nerve cords running on the abdomen form the trunk nervous system. They arise at the subterranean ganglion and run parallel to each other to the rear end of the animal. Each nerve cord has four or five ganglia that correspond to the four leg-bearing segments and perhaps another legless genital segment. The two ganglia of a leg segment are linked by transverse nerve connections.
Sense organs
Many, but not all, tardigrades have point-shaped eyes, either red or black. They are designed as so-called pigment cup oocelli, that is, each eye consists of a cup-shaped pigment cell enclosed by two photoreceptor cells, the concave side of which faces the body surface and thus the light. They are supplied with nerves by the outer, rear lobes of the brain and, in their special form, are only found in tardigrade.
In addition, in some species there are bristle-shaped sensillae on the trunk segments , which presumably react to chemical or contact stimuli. Thread-shaped sensillae, the cirri , are found on the back of the head, especially in many marine species, and are probably organs of touch, while clavae , somewhat thicker and internally hollow threads, are probably chemoreceptors . In many species there are wart-shaped elevations around the mouth opening, the papillae , which probably also have a function in the perception of the environment.
Reproductive organs
Tardigrade basically only have one unpaired gonad (gonad) surrounded by real coelomic tissue, which is located above the digestive tract and attached to the front end of the back body wall by ligaments.
The flagellated sperm are formed in the testicles of the males . Two sperm conductors emanate from it, which unite on the abdomen at the often tubular protruding genital opening, the gonopore, located in front of the anus on the body center line and open to the outside world. The functionally inexplicable duplication of the sperm ladder is seen as an indication of the evolutionary loss of a gonad.
The female ovary, on the other hand, has only one fallopian tube, which, depending on the species, is either to the right or left of the intestine. In the species of one class, the heterotardigrada, its mouth is located in a separate gonopore, usually located in front of the anus, while in the species of the other class, the eutardigrada, it is in the hindgut, which thus becomes a cloaca. Many females have one or two seminal vesicles which, during copulation, take in the males' sperm and store them until they lay eggs.
Hermaphrodite individuals have a gonad known as ovotestis , in which both sperm and egg cells mature and can be released through a shared egg or seminal duct.
distribution
Tardigrades live on all continents worldwide including Antarctica and in all oceans. They can be found in central European rain gutters as well as in regularly iced arctic pools or tropical rainforests , at an altitude of more than 6000 meters in the Himalayan mountains, on remote islands such as the South Sandwich Islands , in the abyssal zone on the floor of the 4690 meters deep Indian Ocean or in the middle of the Atlantic on drifting brown algae . Although they occur in all climatic zones, there is a distribution focus in polar and temperate latitudes.
A biogeographical structure can only be recognized at the family and genus level, which can be associated with the separation of the ancient continent Pangea into Gondwana and Laurasia . A total of ten genera and 22 species are cosmopolitans themselves, that is, to be found all over the world. They are considered remnants of a Pangea fauna that existed before the geological epoch of the Triassic . Most of the other species have a spatially restricted range.
habitat

According to their habitat, tardigrade can basically be described as marine ( marine ), freshwater ( limnic ) or land-living ( terrestrial ), although the separation between the last two categories is only vague, which is why the term limnoterrestrial is often used . All tardigrade, although some of them are highly resistant to dehydration, are dependent on a thin film of water for an active life.
Sea habitat
The marine species live in both salt and brackish water and can be found from the intertidal zone down through the shallow water zone to the abyssal plains of the oceans; at least one species is able to withstand the water pressure that prevails on the floor of the Mariana Trench . If the seabed is muddy, the benthic tardigrades there have mostly adapted to their surroundings through a strong worm-shaped body with shortened extremities; In sandy or pebbly ground that offers crevices and cracks, on the other hand, animals with well-developed stub legs are more likely to be found. One species has apparently specialized in manganese nodules as a habitat, while others in extensive algae carpets or on animals such as stony corals (Scleractinia), bryozoa , mussels (Bivalvia), woodlice (Isopoda), barnacles (Cirripedia), sea urchins (Echinoidea) or Sea cams (Holothuroidea) live partly commensally , i.e. without influencing the host, but partly also parasitically . At least one species lives on floating Sargassum algae in the middle of the open ocean in the Sargasso Sea . Towards the land there is a pronounced sand gap fauna in the top centimeters of the soil on almost all beaches, where the animals live between individual grains of sand. If there are several species, they are mostly distributed over different micro-habitats, which can be separated from one another by differences in humidity or temperature. The transition to the terrestrial forms is formed by those tardigrades that live in marine lichens , which have settled on boulders above the water level usually reached at high tide and are normally only reached by the salty spray.
Freshwater habitat

The limnic species in the true sense of the word live in flowing waters as well as in lakes, ponds, ponds or even individual puddles. Within a lake, tardigrade mostly forms part of the bottom-living sand gap fauna; Finds from a depth of up to 150 meters are known. The animals mostly live in the uppermost, oxygen-containing centimeters of the soil, find themselves towards the lake shore, where loose sand could seriously injure the animals, but mostly a little deeper. Algae and aquatic plants are also colonized, and tardigrades can occasionally be found in water lilies on the surface of the lake .
Often individual individuals can be tracked down in rain gutters; the animals are probably washed by mosses on the roof, so that they are considered terrestrial. Finally, hot springs also form a freshwater habitat that is populated by some species.
Habitat land

The most important terrestrial habitats are moss lawns; the species that live there are known as moss-loving or bryophilic . Because mosses store water in the interstices, however, they can also be viewed as aquatic habitats. Different species are often found in the different zones of the moss; the tardigrade fauna of the rhizoid layer, with which the mosses are rooted in the soil, and the photosynthetically active but dehydration-prone outer layer are not identical. In addition, the animals found in lichens or suitable flowering plants such as bromeliads (Bromeliaceae), pitcher plants (Nepenthaceae) in saxifrage ( Saxifraga ) Mannsschild ( Androsace ) or carding ( Dipsacus ). In all of these plants, water collects in the leaf roots or other cup-shaped parts of the plant; they become a miniature habitat for tardigrade.
Tardigrades are also often found in the leaf litter of forests or in the ground itself, with beech forests apparently being particularly popular. A somewhat more unusual habitat are the glaciers in the high mountains: there, dark deposits of dust or fine-grained rubble can lead to heat absorption during the day and thus to the temporary thawing of the surface; Tardigrades can also be found in the resulting aqueous cryoconite layer. Urban habitats have hardly been studied to date.
Tardigrades are only active when they are at least covered by a thin film of water in their respective substrate, so that they cannot live in arid areas such as deserts . However, many terrestrial species are extremely drought-resistant and can therefore survive regular dry periods and temperature extremes. This fundamental ecological advantage, which they share with some rotifers (Rotifera), they can best play out over competing animals such as roundworms (Nematoda) where the moisture content of the environment is subject to strong fluctuations, which is at least partly the great preference of many tardigrade species Mosses and lichens explained, which dry out in a very short time or can be flooded with water. The colonization of these habitats, to which few other animals have access, is an important reason for the great evolutionary success of the tardigrade.
Population densities
Exact information on population densities is only available for a few species and so far only in random samples: On sandy beaches, up to 35 individuals were found per cubic centimeter of sand, in soils up to 30 individuals per square centimeter of surface were counted, while moss pillows with around 200 individuals per Square centimeters represent a very densely populated habitat as expected. As a rule, however, the population densities are well below this maximum value. As far as it can be said on the basis of the still inadequate data situation, the number of individuals in marine species is usually much smaller than that of land or freshwater forms.
Population densities can be influenced by temperature and humidity, the food supply, the number of parasites and predators or by environmental pollutants, and in the limnoterrestrial species they mostly fluctuate depending on the season, with a first peak in spring and early summer and a second peak in autumn.
Transportation and locomotion
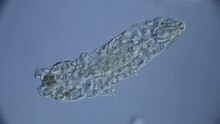
Tardigrade spreads only to a very minor extent through active locomotion; instead, most animals are moved into new potential habitats by wind, water or animals. This passive transport primarily affects the animals' eggs as well as cysts and barrels - life stages that are to a greater or extreme extent independent of environmental extremes.
Marine species can be drifted in ocean currents, which probably help them with special sail-like bulges on their outer skin. Limnoterrestrial species are sometimes transported in the same way by overflowing flowing waters or meltwater. Temporary dry habitats allow eggs or cysts to spread with the wind ( anemochory ), while smaller distances can be bridged in splashes of water. During a storm off Greenland, even adult animals could be detected in falling raindrops, which were probably thrown up by the storm beforehand. Presumably, insects or birds, to which the eggs or cysts stick, also bring tardigrades into new habitats ( zoochories ).
All species need a thin surrounding film of water for active locomotion. They then use the legs of the first three trunk segments, which, unlike the related columbus, can be moved not only in pairs but also individually, for example to scramble over grains of sand or climb around in moss pillows. The claws or adhesive discs on the legs are used to grip the respective substrate. Unlike the first three pairs of legs, the two rear limbs serve to hold on to the ground, and in some species to move backwards; they are adapted to this modified function through the different claw alignment. The highest experimentally measured running speed is 17.7 centimeters per hour; the value makes the Latin name of the group, Tardigrada, derived from the Latin tardus 'slow' and gradus 'step', which means 'slow strider', appear appropriate.
Phototaxis , i.e. the movement to and from light sources, has not yet been adequately investigated. Young animals seem to react negatively photokinetically , that is, they react to light irradiation with faster movements and spontaneous changes of direction, without any specific avoidance reaction being detected. Since light radiation is often associated with thermal radiation and subsequent water evaporation, this behavior is perhaps related to the risk of dehydration, which is more threatening for young animals.
A marine species is able to swim actively; its cuticle is bell-shaped and can expel a directed jet of water by contraction; the animals move like jellyfish according to the recoil principle.
nutrition

Most tardigrades feed on a vegetarian diet, mainly on algal cells that they find either free-living or in lichens. The plant cells of the mosses, on the other hand, rarely belong to the food spectrum, even in the animals that live there permanently. In addition to algae, soil-dwelling species also ingest organic waste along with the bacteria and fungal spores it contains ; but some forms also live completely or partially predatory. Their range of prey includes protozoa , rotifers (Rotifera) and roundworms (Nematoda), but also other tardigrade. Some marine species live as ectoparasites on the skin of sea walnuts or barnacles . On land, tardigrades are poorly adapted to a parasitic way of life because of the constant danger of dehydration; only one species is known here, which may be endoparasitic in land snails .
To feed, tardigrade press their mouth cone against the affected plant cell, the skin of their prey or the body wall of their host. By advancing the needle-sharp stylets, these are then pierced or drilled and the entire cell or body content is sucked out. Particularly large species, on the other hand, can not only take in liquid food, but also suck up their prey as a whole; this particularly affects smaller wheel animals and tardigrade.
Predators, parasites and commensals
Tardigrade's main predators are other tardigrade species, rotifers (Rotatoria) and nematodes, as well as mites (Acari), spiders (Araneae), springtails (Collembola), various insect larvae , but also non-specific "grazing" organisms such as earthworms (Lumbricidae) or various freshwater crabs.
Some tardigrades fall victim to carnivorous fungi , the fine cell threads ( hyphae ) of which are intertwined into loops in which their prey is caught and which then grow into the captured animals. Other fungi such as Harposporium , the asexual stage of the filamentous fungus genus Atricordyceps give conidia, asexual spores, which probably once they are eaten by tardigrades, germinate in the intestine and digest their prey from within. The fungus may also penetrate the cuticle of the animals from the outside. Other important fungal parasites of tardigrade are the potty fungus Sorochytrium milnesiophthora and the yoke fungi Ballocephala sphaerospora and Ballocephala verrucospora .
There are probably a number of parasites among the protozoa ; until that has been studied in detail yet but ciliates Pyxidium tardigradum that preferentially infects tardigrades. It probably lives as a symphoriont , that is, it can only be spread by its host, but does not appear to be dangerous to itself.
Many symbiotic bacteria are often found in marine tardigrade ; The land-living forms also mostly have a rich bacterial intestinal flora. The proteobacterium Xanthomonas campestris , an important plant pest, is believed to be transmitted by tardigrade.
Molting
Tardigrade can only grow in size through regular molting . The non-cellular outer skin (cuticle) together with the stilettos, the lining of the foregut and hindgut and the leg claws are shed. The molt always begins at the front end of the animals, with the fact that the old stilettos and the cuticula of the oral cavity and foregut are expelled. As the body contracts temporarily, it detaches itself from the old skin, which is then stripped off and left behind as an empty shedding shirt ( exuvium ). During this process, which normally takes about 5 to 10 days, the animals are in the simplex stage , without any stylet , in which they cannot ingest any food. During the moulting, new cuticles are formed from the underlying epidermis, while the new stilettos are synthesized in the salivary glands and the claws are built up by special claw glands.
Up to 13 moults in the course of their lives could be detected in individual individuals; Apart from the fact that they make lifelong growth possible, they also serve to reduce internal body pressure, to repel waste materials stored in the cuticle or to remove parasites. Many species lay their eggs in the shedding shirts.
Stages of resistance
Many tardigrade have developed unique adaptations to survive periods of drought, cold spells, sharp fluctuations in the salinity of the water or a lack of oxygen. A contribution to robustness is their Eutelia: After embryonic development, cell division with its sensitive phases hardly takes place (except in the germ line ). This already results in a very high radiation resistance (apart from sterility ), for comparison see the table in → Radiation damage . In addition, some species adapt to the seasons through morphological changes, while others can form thick-walled cysts. The extreme form of adaptation, however, is the so-called cryptobiosis , in which the animals pass into a state close to death in which no more metabolic activity can be registered. All stages of resistance serve to survive adverse environmental conditions and thus give tardigrade an evolutionary competitive advantage over other animal groups. They also play a role in the passive transport of the animals into new potential habitats. The tardigrade is also the first animal known to be able to survive in space.
Cyclomorphosis
Cyclomorphosis is the regular change in body shape in response to cyclical changes in environmental conditions. Most of the life stages of the animals , known as morphs , change from one to the next depending on the season. So far, cyclomorphosis is only known from marine tardigrades from the genera Halobiotus , Amphibolus and Hypsibius . The best studied example so far is found in the species Halobiotus crispae , which lives in the intertidal zone of Greenland : Here, a winter morph called a pseudosimplex can be distinguished from a summer morph . The former is able to withstand temperatures close to freezing and remains mobile even at low temperatures, but it is sterile; only the much less resistant summer morph is fertile.
Cysts
Species that live in freshwater in particular, but also those that colonize mosses or leaf litter, are able to develop stages of resistance known as cysts. To do this, the animals contract to 20 to 50 percent of their previous body size, reduce their metabolism and in some cases also break down their internal organs. This process is accompanied by up to three incomplete, immediately successive moults, at the end of which the animal is surrounded by a multi-walled covering made of cuticle layers. In this condition, the animals can survive for more than a year. As soon as the environmental conditions have changed, the animals can free themselves from their enclosure again within 6 to 48 hours.
Basically, cysts only form in water; they are nowhere near as resistant as the drums mentioned below and, in contrast to these, due to their water content, they are also not heat-resistant.
Anoxybiosis
Anoxybiosis is the term used to describe the tolerance to lack of oxygen. If the concentration of this breathing gas is too low, the osmoregulation breaks down , the animals ingest water and swell significantly. The entire metabolism must now take place anaerobically, toxic waste products accumulate in the body cavity. Most tardigrades are still able to survive such a condition for 3 to 5 days; it is even reported that individual individuals from the genus Echiniscoides were able to survive for up to 6 months in a sealed test tube of decaying barnacles. The return to the normal state takes a few minutes to a few hours, depending on the duration of the anoxybiotic stage.
Anoxybiosis is particularly important in habitats in which the oxygen concentration can temporarily drop sharply, such as in large static water masses. Tardigrade, which lives on carpets of algae in the intertidal zone, are also exposed to an extreme lack of oxygen at low tide and, in evolutionary terms, were perhaps among the first to acquire this adaptation.
Osmobiosis
Osmobiosis is the ability to tolerate fluctuations in the salinity of the water. Tardigrade, like most animals, are only viable if the ion concentration in the water is within certain limits. However, especially those species that live in the intertidal zone can survive astonishingly large fluctuations in the salt concentration for a short period of time. Many species form the barrel described below in very salty water; However, this is a special case of the anhydrobiosis explained below and a direct response to the internal dehydration associated with the high salinity of the environment. The formation of the barrel is therefore not formally regarded as an osmobiotic phenomenon.
Cryobiosis
Cryobiosis is the ability to withstand low temperatures. It is a manifestation of cryptobiosis, which is characterized by an almost total cessation of metabolism. A so-called dormancy condition occurs . The formation of barrels described in more detail below is possible, but not mandatory.
When the temperature drops slowly, the metabolism changes gradually because most of the enzymes lose their activity and instead low-temperature active catalysts become effective, which synthesize the sugar trehalose and other cryoprotective substances: These protect the sensitive biomembrane and replace the essential molecules bound water. Particularly in the extracellular body fluid, so far unidentified, medium-weight molecules that act as freezing germs stimulate the controlled growth of ice crystals, which are stabilized by anti-freezing proteins when they are small.
In this way it is possible for the animals to survive temperatures well below freezing point. This ability allows colonization of the polar regions and high mountain glaciers, where the ambient temperatures regularly drop below freezing point. The subspecies Echiniscus sigismundi groenlandicus , for example, survives 6 to 8 months frozen in winter ice and tolerates regular thawing and freezing in the transition from the tides .
Tardigrades even survived ten days in free space. With the FOTON-M3 satellite , ESA exposed several samples with tardigrade in space to vacuum, cold and UV radiation in 2007. Upon their return, the scientists found survivors even among those tardigrade exposed to the most extreme conditions.
Because tardigrades can survive extreme temperatures of −273 degrees Celsius if they cool down sufficiently slowly, it has been suspected that they are extraterrestrial life forms . Such temperatures were never even remotely reached in their natural habitat, and the ability thus could not have arisen on earth through natural selection . This view is countered by the fact that a living being that survives on earth below freezing temperatures by replacing water in its tissue with protective substances such as trehalose and controlling the freezing process itself can potentially survive temperatures of −273 degrees Celsius without further evolutionary adaptations. Tardigrade are multicellular polyextremophiles .
Anhydrobiosis
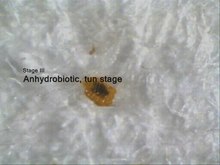
Anhydrobiosis using Richtersius coronifer as an example .
Phase 1: still hydrated and active when dehydrated.
Phase 2: after 10 minutes the legs have dried out and pulled in.
Phase 3: barrel stage (anhydrobiosis).
Phase 4: restitution after addition of water.
Anhydrobiosis is the ability to survive dehydration of the body through heavy water loss. It is found in almost all terrestrial species and is associated with the formation of cylindrical, immobile resistance stages, the barrels . Because no more metabolism is detectable in the anhydrobiotic state in barrels, the anhydrobiosis also falls under the generic term cryptobiosis.
In some species, numerous individuals gather before the actual barrel formation and form a loose ball. This aggregation is seen as a behavioral adaptation and probably shields the animals inside from environmental influences a little more, so that the dehydration process is a little slower for them - too fast dehydration can also lead to death in tardigrade.
The barrel formation begins with morphological reactions: the legs are drawn in and the overall body surface is greatly reduced. Lipids , fat-soluble substances, which, among other things, are supposed to protect against fungal attacks, are increasingly released through pores in the cuticle . As a result of a phase change in these lipids, the cuticle suddenly becomes impermeable to water at a specific point in time. This reduces water evaporation and allows the preparation time to be extended to the anhydrobiotic state, during which the protective compounds must be synthesized.
The main problem with anhydrobiosis is that the structural integrity of important macromolecules such as proteins , phospholipids or nucleic acids must be preserved at all costs, otherwise irreversible damage would result in the cells. Most of these connections are surrounded by loosely attached water molecules, the loss of which would trigger uncontrolled reactions between them. The solution to the problem that the tardigrade found in the course of their evolutionary history consists in replacing the bound water during dehydration with other compounds that can easily be broken down again during rehydration (wetting by water). The most important of these compounds is the sugar trehalose , which is produced in large quantities during preparation for the anhydrobiotic state and is sometimes enriched by a factor of 23 compared to the initial state. It not only protects the biomembranes and prevents reactions between the dehydrated proteins and other cell components such as carbohydrates, but also prevents uncontrolled oxidation , which could also destroy important macromolecules. New research suggests that tardigrades have a new type of disordered protein that, when dehydrated, is increasingly produced and vitrified , that is, form a glass-like, protective material. In addition to this family of proteins and trehalose, glycerine probably also plays a role in these processes. Lipid droplets meanwhile have the task of keeping body tissues separated from one another, which in the normal state do not run the risk of getting too close, but in the barrel state they come into contact and could trigger undesired cross-linking reactions. It takes about 5 to 7 hours for the animals to return from their normal state to a stable barrel stage.
As soon as the animals have reached the anhydrobiotic state, at least in a dry environment no more metabolic activity can be detected; in particular, the consumption of oxygen falls to zero. At this stage, the animals can survive extreme dehydration, immersion in brine, ether , pure ethanol or liquid helium , temperatures between −196 ° Celsius and +90 ° Celsius, high hydrostatic pressure and vacuum .
As soon as there is enough water again, the transformation back to the normal state, the restitution , takes place. To do this, the trehalose is broken down aerobically, i.e. using atmospheric oxygen, and then replaced by water. The recovery time depends on the temperature, the oxygen content and the pH value of the environment as well as the time spent in the cryptobiotic state and also varies somewhat from species to species. In most cases it is between ten minutes and several hours, in the case of a very long barrel condition even a few days.
Whether the return to an active stage of life is successful depends primarily on the correct sequence of barrel formation and the nutritional status of the animals. Particularly important in this context are the lipid reserves, which not only play a role in the cryptobiotic state, as mentioned above, but can also be converted into glycerol and trehalose and also provide energy for the restitution process. As long as the animals have the opportunity to replenish these reserves at regular intervals and, moreover, any dehydration takes place only slowly, they can switch from the active to the anhydrobiotic state and vice versa at regular intervals. For example, individuals of the genus Echiniscoides who live in the intertidal zone can be experimentally put into a cycle with a six-hour period.
Tardigrades in the cryptobiotic state raise questions about the definition of life : metabolism, growth and reproduction are often cited as characteristic features of a living organism. However, none of these properties are found in the barrel stage, so that the return to the active state could be described as "resurrection from the dead" in the words of Lazzaro Spallanzani, an Italian naturalist of the 18th century. On the other hand, many, but not all, tardigrade return from the anhydrobiotic stage to an active state, which led the tardigradologist John H. Crowe to ask in 1975:
- Does this then mean that they "died" while they were "dead"?
- Does that mean, then, that they "died" while they were "dead"?
In Crowe's view, the definition of life cannot be reduced to single features; Instead, he sees the continued existence of the structural continuity and integrity of a living system as being "vital" in the literal sense of the word.
Reproduction and life cycle
Tardigrades can reproduce asexually as well as sexually. Most species reproduce exclusively sexually.
Asexual reproduction
Asexual reproduction is only possible through parthenogenesis , i.e. the maturation of unfertilized eggs, which subsequently develop into females. It occurs in a number of species, including the genera Echiniscus and Pseudoechiniscus . Male animals are often not known, even if dwarf males could be described afterwards in some cases. In these and other species a coexistence of parthenogenetic and sexual reproduction can be determined; the males are basically able to distinguish suitable, sexually active, from unsuitable, parthenogenetically reproducing females. An important advantage associated with parthenogenesis is the lack of effort in finding a partner, a serious disadvantage is the reduction in genetic diversity. The negative consequences of this, however, are partially offset by the ability to simply avoid adverse environmental influences through cryptobiosis.
Sexual reproduction
Sexual reproduction in tardigrade mostly, but not always, causes the existence of two separate sexes. Some hermaphroditic (hermaphrodite) species are known to be capable of self-fertilization; Egg and sperm cells mature in the same gonad. They too have the advantage of not having to spend any energy looking for a partner, but differ from parthenogenic individuals who ultimately clone themselves through the possibility of genetic recombination .
For the vast majority of species, however, there are separate sexes, which, however, cannot always be easily distinguished from the outside. Where there are detailed studies of the population structure, there is almost always a clear excess of females.
The fertilization can take place outside as well as inside the body of the female, the oviposition for the female is mostly connected with a molt. The details of both processes depend, among other things, on the animal's habitat.
In marine species, the males' sperm are usually deposited in the seminal vesicles of the females, which then lay their eggs freely and attach to the substrate such as algae cells. In addition, it also happens that the female lays her unfertilized eggs immediately after molting in the rejected cuticle, where they are then fertilized externally by the male.
In limnoterrestrial species, the sperm are occasionally injected into the gap between the old and new skin before or during the molting of the female; as soon as the female lays her eggs in the discarded moulting shirt, fertilization takes place. Especially with terrestrial species there is often copulation, for which the male clings to a female; Such a connection is not given up even in the case of external disturbances of the sexual act. The male now introduces his sperm into the female's genital tract before the female has finished moulting. Alternatively, the sperm can also be deposited in seminal vesicles or injected directly through the outer skin into the body cavity; in the latter case fertilization takes place in the female gonad.
The number of eggs laid varies between 1 and 35 depending on the species, with a steady increase in this number with increasing age. Over a lifetime, single females can produce over 100 eggs. In marine species they are thin or thick-shelled depending on the environmental conditions, while different egg shapes can be observed in terrestrial species: Smooth eggs are mostly deposited in the moulting shirts of the females, while eggs with thick, elaborately ornamented and probably dehydration-resistant shells are free on moss plants or Pieces of bark are glued on. Some freshwater species use the shed exoskeletons of insects or other arthropods as a depository.
Special behavioral adaptations during reproduction are only found in rare cases: A primitive “mating ritual” has been observed in some species. The male strokes his female with the cirri on the head. After some time, stimulated in this way, the female lays her eggs on a grain of sand, on which the male then deposits his sperm. The fact that some females drag the shedding shirts in which they have laid their eggs for a while is sometimes interpreted as a simple case of "brood care".
Under favorable conditions, the young hatch after about 5 to 40 days. To do this, they absorb liquid from the environment until their expanding body bursts the egg shell. Some species also use their mouth stylets to break open the egg shell. The entire process usually only takes a few minutes.
The young animals are mostly undyed and have fewer bristles, cirri or claws than the adults. Otherwise they are very similar to these, so that their development takes place directly, i.e. without the larval stage. In some species from the class Heterotardigrada, the anus and the genital opening form only after one or two moults; this is sometimes interpreted as an indication of an indirect development. In most cases the young animals only grow through an increase in the individual cell volume, but not through an increase in the number of cells, so that many tissues already have the final number of cells of the adult form after hatching. However, even in later stages, mitoses (cell division) can occasionally be detected, which presumably serve the purpose of replacing dead cells. In any case, sexual maturity is only reached after several moults.
The normal lifespan of tardigrade is between three months and two and a half years; it corresponds to the actual lifespan of most marine species. In the limnoterrestrial species, however, the life of the animals is sometimes or often interrupted by cryptobiotic states, during which the affected individuals do not age. In this way, moss-living species often achieve a real lifespan of four years or more; in individual cases they can last for decades. A tardigrade was discovered in the dried out moss of a botanical museum, which after 120 years in an anhydrobiotic state could be "revived".
Tardigrade and man
Tardigrade can be found in almost all human habitats, but due to their small size and unusual way of life, they are hardly noticeable. Since they have no immediate economic, medical or veterinary importance, they are unknown to most people. Those who see them for the first time often describe them as “cute”, an adjective that can be found in reputable zoological publications and is probably inspired not only by the bear-like body shape, but also by the clumsy way the animals move. Keeping them in a monoxenic culture, i.e. together with at least one other species, is possible.
Information on the hazard is not available; however, one species, Thermozodium esakii , may have become extinct. Since tardigrade is very sensitive to environmental toxins , especially sulfur trioxide , there are ideas to use the animals as indicators of the environmental quality of a location. Studies of damage to Tardigrada from heavy metal pollution from mosses were presented by Hungarian researchers. Studies on urban habitats are not yet available.
Tribal history
Modern forms
| Representatives of related taxa |
|---|


.jpg)
The closest relatives of the tardigrade can be found in two very different groups: the arthropods , which include crustaceans (Crustacea), arachnids (Arachnida), millipedes (Myriapoda) and insects (Insecta), are the most extensive animal phyla, while the columbus (Onychophora), which can be described as worms with legs, form a rather obscure taxon. Tardigrades, arthropods, and stumpworts very likely form a natural family group together, a so-called monophyletic taxon , which is referred to as Panarthropoda. The segmentation of the body occurring in all animals of this group as well as the appearance of paired body appendages can be viewed as a common derived characteristic; the structure of the cuticle is also shared by all three taxa.
Within the Panarthropoda, tardigrades are traditionally grouped together with the stumpefoots as protoarthropods:
| Panarthropoda |
|
||||||||||||
|
|
Originally, the idea played a role that columbus and tardigrade have not yet reached the full organizational level of arthropods. In evolutionary terms, however, the distinction between more or less highly developed forms is now considered out of date; modern classifications should only reflect the actual phylogenetic relationships of the taxa to one another.
The common features of the Protoarthropoda are the sac-like body appendages, the stump legs, which can be found in both columbus and tardigrade. There are also a number of other homologies , i.e. features that can be traced back to a common precursor structure. In particular, the tardigrade head, which is believed to be made up of three segments, very likely corresponds to the first three units of the colobus body. Their stilettos can accordingly be viewed as strongly modified body appendages of the second segment and would thus be homologous to the jaws of the velvet worms. The claw structure is also very similar in tardigrade and columbus. Despite these striking similarities, it is controversial whether the Protoarthropoda form a natural family group, since presumably all of the cited features are symplesiomorphies , that is, they were already found in the ancestors of all Panarthropoda. At the beginning of the 21st century, a preference for a sister-group relationship between tardigrade and arthropod emerged instead:
| Panarthropoda |
|
||||||||||||
|
|
The further relationship of the tardigrade is traditionally seen in the annelids (Annelida). With them they share, among other things, soft, non-articulated body appendages with final hardened “claws” and a terminal mouth position that closes the body towards the front. This Articulata hypothesis, which goes back to the French naturalist Georges Cuvier , still had many supporters at the beginning of the 21st century , but was increasingly questioned towards the end of the 1990s due to morphological and molecular genetic findings. It was replaced by a concept that in a group of wormlike animals, Cycloneuralia, provides the closest relatives of Panarthropoda: At this taxon counting thread (Nematoda) and string worms (Nematomorpha), but also three rather obscure animal groups priapulida (Priapulida ), Hookweed (Kinorhyncha) and corset animals (Loricifera). Like the Panarthropoda, all of these taxa are characterized by the fact that they reject their non-cellular outer skin or cuticle, at least during individual stages of their life cycle; they are therefore called molting animals (Ecdysozoa):
| Molting animals (Ecdysozoa) |
|
||||||
|
|
Extinct forms
Tardigrade fossils are extremely rare and therefore contribute little to understanding the development of this group of animals. In addition to some finds from the early geological period , only a few individuals from the late geological period are known to have been preserved in amber .
The lobopods , a group of worm-like animals from the geological epochs of the Cambrian and Ordovician , which, however, probably had their origins in the previous Ediacarian , are increasingly seen as possible representatives of the Tactopoda, the taxon of arthropods and tardigrade . Similar to the columbus, they ran on non-articulated, bag-like stub legs and are therefore traditionally assigned to this animal tribe. Some modern cladistic studies consider this assessment to be unfounded and instead see the lobopods as a non-natural relational group from which both the velvet worms and the tactopods arose. A suggested variant that expresses this alternative view is shown in the following diagram:
| Panarthropoda |
|
|||||||||||||||||||||||||||||||||||||||
|
|
The genus Aysheaia , which still has many original features such as a simple row of unspecialized long legs or a fine superficial ringing that does not match the inconspicuous body segmentation, thus forms the evolutionary sister group of all other Panarthropoda, which then divides into the stumpworts and all other groups split up. A number of lobopods such as the heavily armored genera Cardiodictyon , Hallucigenia or Paucipodia presumably form an extinct side branch, which faces the unassigned genus Luolishania on the one hand and an unnamed taxon from the species Kerygmachela kierkegaardi and the Tactopoda on the other.
Kerygmachela kierkegaardi , according to this hypothesis the immediate sister group of the Tactopoda, is known from the early Cambrian of Northern Greenland and, like tardigrade and arthropod, is clearly segmented on the outside. It is unclear whether the mysterious Anomalocaris fossils also belong to the trunk line of the Tactopoda or, as shown above, are more closely related to the arthropods than to the tardigrade.
The first fossils that can be clearly assigned to the tardigrade come from the Siberian Kuonamka Formation. In the 530 million year old limestone from the Middle Cambrian, four individuals have been preserved there, which can be identified as tardigrades based on their body shape and size. The still undescribed species apparently had three, maybe even four pairs of legs, which ended in a pair of unequal claws, and had a cuticle structure that was very similar to that of today's animals. The animals , which are already very specialized, can possibly even be assigned to one of the modern classes, the heterotardigrada .
How the transition from the robust lobopod to the sub-millimeter-sized tardigrade took place is unknown. A possible mechanism would be progenesis , a process in which the gonads form prematurely during embryonic development and therefore sexual maturity occurs earlier than the initial state. This now arrests the further development and differentiation of the body, so that larval or juvenile features such as a significantly small size appear in the adult animal, a phenomenon that is referred to as pedomorphosis .
Even when tardigrade first conquered terrestrial habitats cannot be said with certainty due to the lack of fossil records. Since tardigrade can often be found in moss pillows today, mosses were very likely the first plants to colonize the country and were exposed to similar challenges as the tardigrade themselves in terms of the risk of dehydration, it is very possible that mosses and tardigrades stepped ashore together with the necessary adjustments.
At this point at the latest, the animals must have developed the unique adaptation to periods of drought, known as cryptobiosis. It allowed them to simply avoid adverse environmental conditions by temporarily switching off their metabolism. This has the consequence that at least the environmental selection is only relatively weak, and thus explains the very slow, Bradytelian rate of evolution of the physical appearance, the phenotype .
It is therefore not surprising that the later tardigrade fossils obtained from the Cretaceous period hardly show any change in body shape compared to today's species. Beorn leggi, for example, a species obtained from Canadian amber , can already be assigned to the Eutardigrada class; a very poorly preserved unnamed young animal from the same locality possibly belongs to the class Heterotardigrada. Further amber fossils are known from the US state of New Jersey , which presumably originate from the late Cretaceous period called the Turonian . They, too, are presumably assigned to the Eutardigrada and do not allow any further conclusions than that this line of development has existed for more than 65 million years.
The only other tardigrade fossils are eggs a little over 7,000 years old that have survived in sub-Antarctic peat bogs . Since different species of tardigrade have different temperature and humidity preferences, there are considerations to use the eggs that appear in the different layers of bog, similar to plant pollen, to determine the climate at that time.
Systematics
There is no serious doubt that tardigrades form a monophyletic taxon , that is, go back to a common parent species and include all descendants of this species. Important synapomorphies , common, derived features, are the telescopically retractable legs and the mouth stylet.
By 2005, around 930 species were described, including 160 marine forms. The actual number of species is naturally unknown, but is estimated at around 10,000. In addition, many “species” that are delimited according to morphological criteria are presumably concealed instead by groups of cryptic species that can only be distinguished by molecular genetics, which would further increase the biodiversity of the taxon.
There are three different classes, the relationships between which are still unclear:
- As heterotardigrada is called the "armored" tardigrades even if not all species actually have a back and hardened into individual armor plates (sclerites) divided cuticle. In many forms, conspicuous appendages like cirri and clavae can be observed; the legs can end in both claws and adhesive discs. The genital opening is always directly on the body surface, Malpighian glands for excretion and osmoregulation do not occur. Heterotardigrada are found in both marine and limnoterrestrial habitats.
- As eutardigrade one calls the "naked" water bears, their skin is thin and does not harden. Noticeable sensory hairs on the head are never found in this group; the legs basically end in claws. In contrast to the heterotardigrada, the fallopian tube joins the rectum, which becomes a cloaca; the specialized Malpighian glands serve for excretion. Most of the Eutardigrada live in freshwater or on land, although some marine species also exist.
- The Mesotardigrada are only known from a single, lost species, Thermozodium esakii , which is formally placed in a family Thermozodiidae. According to the description, it has a lateral cirrus on each head, but no clavae; the mouth is surrounded by four wart-like protrusions (papillae). Papillae are also located at the base of the leg, while the end of the leg merges into 6 to 10 simple claws; Malpighian glands are present. The species was found on algae pads in a hot spring, the type -locality, near the Japanese city of Nagasaki , but it was destroyed by an earthquake after the Second World War. Since no type material has been preserved and the species has not yet been found again, its existence is now considered dubious and its name is accordingly the noun dubium .
It is often assumed that one order of the Heterotardigrada, the predominantly marine Arthrotardigrada, includes the most primitive tardigrade species, from which the forms of the other Heterotardigrada order Echiniscoidea and in particular the limnotardigrada family Echiniscidae have developed, before these in turn the other tardigrade class, Eutardigrada whose species are mainly found in freshwater and on land:
| Tardigrade |
|
||||||||||||||||||||||||
|
|
However, this hypothesis has not yet been confirmed; preliminary molecular genetic data suggest that both hetero- and eutardigrada are naturally related:
| Tardigrade |
|
||||||||||||
|
|
Research history
Tardigradology, or tardigrade research, dates back to the 18th century . The German naturalist Johann Conrad Eichhorn was probably the first person to observe the animals on June 10, 1767. Since he published his discovery in his work Contributions to the natural history of the smallest aquatic animals that cannot be seen with the naked eye and that are in the waters in and around Gdansk only in 1775 and thus years later, today the Quedlinburg pastor Johann August Ephraim Goeze is mostly valid as the discoverer of the group. He could make claims to his first observation on 10 December 1772 published his description of the animals in a self-written notes of translated by him from French and published in 1773 magazine Mr. Karl Bonnets treatises from the Insektologie . There he wrote, among other things:
“This little animal is strange because the whole structure of its body is extraordinary and strange, and because in its outward form, at first sight, it has the greatest resemblance to a small bear. This also motivated me to give him the name of the little water bear. […] However, one should not be afraid to contemplate these predators of the invisible world. "
Just one year later, the return from the anhydrobiotic state was observed for the first time, which the Italian naturalist Lazzaro Spallanzani shortly afterwards described as "resurrection from the dead". It was also he who named the group Il Tardigrada in the second volume of his book Opuscoli di Fisica animale e vegetabile , published in Modena in 1776 . In 1790 the animals were included in the work Systema Naturae by the Swedish naturalist and systematist Carl von Linné .
The first scientific monograph appeared in 1840, nine years later the first marine tardigrade was discovered; the scientific description of the first fossil species, Beorn leggi , however, had to wait more than a century until 1964.
The systematic position of the animals was unclear from the start; during the whole 19th century, they were taxonomically optionally with the rotifers (Rotifera) or sea spiders grouped (Pycnogonida) or to the arthropods provided (Arthropoda). Even in the first half of the 20th century , their exact phylogenetic relationships remained controversial, so that finally the Italian tardigradologist Giuseppe Ramazzotti elevated the group to the rank of its own animal tribe. The further classification of the taxon goes back to the German biologist Ernst Marcus, who in 1929 divided all tardigrades into the subgroups of hetero- and eutardigrada; In 1937, Gilbert Rahm introduced the Mesotardigrada as the third group.
Miscellaneous
- The unsuccessful landing of the Israeli lunar lander Beresheet scattered several thousand tardigrade. Since they can do without oxygen and can be brought back to life after warming up from the frozen state, they were selected for this moon mission. “Like dead material, they can survive for years when dried out. They should also have survived the crash on the moon, ”Nova Spivack assured the US magazine Wired . Nova Spivack is the director of the “Arch Mission” foundation.
literature
- EE Ruppert, RS Fox, RD Barnes: Invertebrate Zoology. A functional evolutionary approach. 7th edition. Brooks / Cole, London 2004, ISBN 0-03-025982-7 , p. 510.
- RC Brusca, GJ Brusca: Invertebrates. 2nd Edition. Sinauer Associates, Sunderland Mass 2002, ISBN 0-87893-097-3 , p. 469.
- IM Kinchin: The biology of tardigrades. Portland Press, London 1994, ISBN 1-85578-043-7 , p. 186.
- M. Blaxter, B. Elsworth, J. Daub: DNA taxonomy of a neglected animal phylum: an unexpected diversity of tardigrades. In: Proceedings of the Royal Society of London. Series B. London 271.2004, p. 189. ISSN 0375-0434
- GE Budd: The morphology of "Opabinia regalis" and the reconstruction of the arthropod stem group. In: Lethaia. Taylor & Francis, Oslo 29.1996, ISSN 0024-1164 , p. 1.
- RA Dewel, WC Dewel: The place of tardigrades in arthropod evolution. In: RA Fortey, RH Thomas (Eds.): Arthropod Relationships. (= The Systematics Association Spec. Volume 55). Chapmann and Hall, London 1998, ISBN 0-412-75420-7 , p. 109.
- RA Dewel, DR Nelson, WC Dewel: The brain of "Echiniscus veridissimus" Peterfi, 1956 (Heterotardigrada). A key to understanding the phylogenetic position and the evolution of the arthropod head. In: Zoological Journal of the Linnean Society. Blackwell, Oxford 116.1996, ISSN 1096-3642 , p. 35.
- JR Garey, DR Nelson, LY Mackey, L. Li: Tardigrade phylogeny, Congruency of morphological and molecular evidence. In: Zoologischer Anzeiger. Elsevier, Jena 238.1999, ISSN 0044-5231 , p. 205.
- A. Hejnol, R. Schnabel: The eutardigrade Thulinia stephaniae has an indeterminate development and the potential to regulate early blastomere ablations. In: Development. 132.2005, p. 1349.
- A. Jörgensen, R. Kristensen: Molecular Phylogeny of Tardigrada - investigation of the monophyly of Heterotardigrada. In: Molecular Phylogenetics and evolution. Elsevier, Amsterdam 32.2004, 2, ISSN 1055-7903 , p. 666.
- DR Nelson: Tardigrada. In: JH Thorp, AP Covich (Ed.): Ecology and Classification of North American Freshwater Invertebrates. Academic Press, San Diego Ca 2001, ISBN 0-12-690647-5 , p. 527.
- DR Nelson: Current Status of the Tardigrade. Evolution and Ecology. In: Integrative and Comparative Biology. Lawrence 42.2002, ISSN 1540-7063 , p. 652.
- DR Nelson, NJ Marley: The biology and ecology of lotic Tardigrada. In: Freshwater Biology. Blackwell, Oxford 44.2000, ISSN 0046-5070 , p. 93.
- Joh. August Ephraim Goeze: Mr. Karl Bonnet's treatises from insectology. (PDF; 32.7 MB). Supplement by the translator JAE Goeze, Halle 1773. (Original text: This little animal is strange because the entire structure of its body is extraordinary and strange, and because its outward form, at first sight, looks most like a small bear has. pp. 367–375)
Web links
- Tardigrada Astronomy Picture of the Day of NASA from March 6, 2013
- Tardigrada Newsletter (Engl.)
- Tardigrades: photos and videos (Engl.)
- Extensive factual texts, photo galleries and short films on tardigrade
- Taxonomy down to the genus level
- Microscopy UK
- Research with tardigrade in space
- Swiss research center for the ecology, physiology and evolutionary biology of tardigrades
- A recent claim that tardigrades got a sixth of their DNA from microbes is starting to unravel (engl.)
- Claudia Ruby: Tardigrade - The cuddly survivor Bavaria 2 radio knowledge . Broadcast on July 25, 2019 (podcast)
- But not indestructible - that is the weak point of tardigrade
Individual evidence
- ↑ Andreas Hejnol, Ralf Schnabel: The eutardigrade Thulinia stephaniae has an indeterminate development and the potential to regulate early blastomere ablations. In: Development. 132, 2005, pp. 1349-1361. doi: 10.1242 / dev.01701
- ↑ Daiki Horikawa et al .: Radiation tolerance in the tardigrade Milagnes tardigradum . Int. J. Radiat. Biol. 82, 2006, doi: 10.1080 / 09553000600972956 ( free full text ).
- ↑ Emma Brennand: Tardigrades: Water bears in space . Ed .: BBC. May 17, 2011 ( bbc.co.uk [accessed November 30, 2019]).
- ↑ Peter Reuell: Harvard study suggests asteroids might play key role in spreading life. In: Harvard Gazette. July 8, 2019, Retrieved November 30, 2019 (American English).
- ↑ Lorena Rebecchi et al .: Resistance of the anhydrobiotic eutardigrade Paramacrobiotus richtersi to space flight (LIFE-TARSE mission on FOTON-M3) . Journal of Zoological Systematics and Evolutionary Research 49, 2011, doi: 10.1111 / j.1439-0469.2010.00606.x ( free full text ).
- ^ Neil A. Campbell et al.: Biology. 8th, updated edition. 2011, ISBN 978-3-8273-7287-1 .
- ↑ D. Schulze-Makuch et al .: Tardigrades: An Example of Multicellular Extremophiles. In: Joseph Seckbach et al.: Polyextremophiles - life under multiple forms of stress. Springer, Dordrecht 2013, ISBN 978-94-007-6487-3 , pp. 597-607.
- ↑ Thomas C. Boothby, Hugo Tapia, Alexandra H. Brozena, Samantha Piszkiewicz, Austin E. Smith: Tardigrades Use Intrinsically Disordered Proteins to Survive Desiccation . In: Molecular Cell . tape 65 , no. 6 , March 16, 2017, ISSN 1097-2765 , p. 975-984.e5 , doi : 10.1016 / j.molcel.2017.02.018 , PMID 28306513 ( cell.com [accessed June 20, 2017]).
- ↑ DD Horikawa, T. Kunieda u. a .: Establishment of a rearing system of the extremotolerant tardigrade Ramazzottius varieornatus: a new model animal for astrobiology. In: Astrobiology. Volume 8, Number 3, June 2008, pp. 549-556, doi : 10.1089 / ast.2007.0139 , PMID 18554084 .
- ↑ Fumihisa Ono et al .: Effect of ultra-high pressure on small animals, tardigrades and Artemia. Cogent Physics 3, 2016, doi: 10.1080 / 23311940.2016.1167575 (free full text).
- ↑ B. Vargha, E. Otvös, Z. Tuba: Investigations on ecological effects of heavy metal pollution in Hungary by moss-dwelling water bears (Tardigrada), as bioindicators. In: Ann Agric Environ Med. 2002; 9 (2), pp. 141-146. PMID 12498580
- ↑ Klaus J. Müller, Dieter Waloszek, Arcady Zakharov: 'Orsten' type phosphatized soft-integument preservation and a new record from the Middle Cambrian Kuonamka Formation in Siberia. In: New Yearbook for Geology and Paleontology. Papers 197 (1), 1995, pp. 101-118.
- ↑ JAE Goeze: Mr. Karl Bonnets treatises from the insectology. JJ Gebauer, Halle, 1773. Digitized version accessed September 9, 2016. Quoted from www.baertierchen.de .
- ↑ Patrick Illinger: Tardigrades are the first inhabitants of the moon . In: Süddeutsche Zeitung , August 7, 2019