
blossom

1. flower base (Receptakulum)
2. sepals (sepals)
3 petals (petals)
4. stamens (stamens)
5. stamp (pestle)

The flowering of a plant is - in a broad definition - an unbranched short shoot with limited growth that can only be observed after the flowering stage has reached maturity , the leaves of which are indirectly or directly in the service of sexual reproduction : indirectly as protective or attracting organs ( flower envelope ), directly through formation the reproductive organs ( stamens and carpels ).
A narrower definition limits the term flower to the angiosperms . An angiosperm flower consists of the following components (but not all of them have to be represented): the perianth as a perigone or separated into a calyx and petal envelope , the androeceum from stamens, the gynoeceum from carpels and the flower axis ( flower base ). The stamens form the pollen , which gets onto the stigma of the carpels during pollination , germinates there and in the course of fertilization fertilizes the ovules in the carpel, more precisely the ovum located in the ovule. This develops into an embryo , the ovule develops into a seed and the flower into a fruit .
Flowers that only contain carpels are colloquially referred to as "female", those that contain only stamens are referred to as "male" and flowers that have both stamens and carpels are referred to as bisexual or hermaphrodite flowers (see below: Sexuality ).
Definitions

The broad definition of blossom (from Middle High German / Old High German bluot with the plural blüete ), as it often occurs in the botanical specialist literature, partly explicitly or implicitly includes the naked ones . Only rarely is it implicitly restricted to the angiosperms by naming the parts in the definition. In the Strasburger textbook of botany , too , the flower is assigned to all seed plants, albeit with restrictions: “In spermatophytina, the sporophylls are almost always on short shoots with limited growth. Such [...] structures can be called flowers, even if this term is mostly only used for the primarily hermaphrodite sporophyll levels of angiosperms and possibly closely related fossil seed plants. ”The lexicon of biology also briefly defines the flower as“ a short sprout with limited growth who carries the sporophyll ".
Fossil representatives outside the seed plants that formed flowers are the Bennettitales and Caytoniales . The definition of the flower also corresponds to the sporophyll levels of the bear moss and horsetail , but these are not dealt with in more detail here.

In English-speaking countries, blossom (botanical: flower ) is usually more narrowly defined as angiosperm blossom, e.g. B .: "Shortened, hermaphroditic reproductive stem axis with limited growth, which consists of megasporangia (carpels) and microsporangia (stamens) and a sterile flower envelope made of at least one sterile, flat organ."
morphology
In angiosperms, the individual flower organs are arranged within the flower in a fixed order from bottom to top, or, since the flower axis is compressed, from outside to inside. In this order they are also applied ontogenetically (in the course of flower formation).
Perianth
The perianth (perianth) are sterile sheets , but that clearly belong to bloom. An inflorescence is only formed by Gnetopsida and angiosperms . If all the bloom cladding sheets are uniformly designed (Homoiochlamydeisch), the blooming envelope is called perigon or simple perianth , the leaves as tepals (for example with the tulip ). The perigone can consist of one circle (haplo- or monochlamydeic), two or more circles or several screw circuits; multiple perigone . A double perianth (di-, heterochlamydeic flowers) consists of dissimilar bracts. The outer, mostly green, are the sepals (sepals) and form the calyx, the inner, often strikingly colored leaves are the petals and form the crown (corolla). The evolutionary origin of the flower envelope is interpreted from bracts or stamens , depending on the kinship group .
Microsporophylls


Microsporophylls are leaves that bear sporangia. The male microspores are formed in the sporangia (pollen sacs) . The microsporophylls are developed very differently depending on the seed plant group, and a homology to the leaves (φύλλον phýllon "leaf") has not been fully clarified. They could therefore also be called microsporangia carriers more neutral. In ginkgo , the microsporophyll is a stem with two pollen sacs hanging from its tip; Too many, the microsporophylls are screwed on the flower axis. In conifers , the male flower is cone-like and consists of many microsporophylls, usually arranged in a screw-like arrangement; there are two to 20 pollen sacs on each microsporophyll. In the cycads (cycads) which are imbricated and microsporophylls carry five to 1000 pollen sacs; the microsporophylls are screw-shaped in the male flower. In the Welwitschia , the six microsporophylls that have grown together carry three pollen sacs each. In Gnetum the only microsporophyll has one or two terminal pollen sacs. In ephedra , the microsporophyll is often bifurcated and carries two to eight groups of mostly two intergrown pollen sacs.
The microsporophylls of angiosperms are called stamens (stamina). A stamen is usually in the filament (filament) and the dust bag divided (anther). The latter consists of two counters, each with two pollen sacks, connected by a connector. The entirety of the stamens is called Androeceum . The number of stamens per flower is between one and around 2000 pieces.
Megasporophylls

The megasporophylls carry the ovules characteristic of seed plants , the structures in which the female reproductive organs are formed. They too could be described more neutrally as carriers of megasporangia. Only the megasporophylls of the angiosperms are called the carpel .
The ovules of the cycads are clearly on leaves, they are phyllospor. The scale-like or shield-shaped megasporophyll is clearly stalked and has two ovules at the lower edge of the blade , in Cycas there are up to eight ovules along the rachis . The conifers have one to 20 ovules on the surface of a flat seed scale. The seed scale is a modified short shoot. The ovules in conifers are rarely at the end of short shoots, as in the yew tree ( Taxus ). Ginkgo forms two ovules at the tip of a forked stem. The Gnetopsida form only one terminal ovule per flower.
In angiosperms the ovules in the are Carpel (carpel) included, hence the name "angiosperms". The totality of the carpels forms the gynoeceum . The number of carpels per flower is between one and around 2000. Usually, a carpel is divided into a stalk zone and a tubular zone, the hollow area in which the ovaries lie (also called the ovary). The ovules attach to placentas . Above this is often a stylus on which the scar is located, the surface for conceiving the pollen. The scar is connected to the ovules by a transmission channel or tissue. The carpels can be free (apocarpes or chorikarpes gynoeceum) or fused together (coenocarp). An overgrown gynoeceum is called a stamp (pestle). Sterile flowers have at most reduced stamens and carpels and often have the function of a display device .
Flower axis

The petals attach to the flower axis (flower base or receptaculum). It can be compressed, stretched and shaped differently. Single or multiple whorls can be raised by intercalary meristems : gynoeceum ( gynoceum , for example in the caper bush ), gynoeceum and androeceum ( androgynophore , for example in passion flowers ), or gynoeceum, androeceum and crown ( anthophore , many carnation plants ).
Annular areas around the center can also be raised. One example is a stamen-Corollentubus as many composite flowers is typical, or the cup-Kronbecher many cucurbits . A widened flower axis is called a flower cup (hypanthium). If all parts except the ovary are lifted up, a flower tube arises, which carries the flower envelope and the stamens. The result is a perigynous flower cup. The ovary is then referred to as central. An upper ovary stands freely on the flower axis. If the outer sides of the carpels are included in a flower cup, a subordinate ovary is created, which means it stands below the flower envelope.
Nectaries
Nectaries are glands that produce nectar as a lure for the pollinators. This usually consists of a sugar solution , which can also contain proteins , amino acids and vitamins . Oils are formed less often (for example in the loosestrife , Lysimachia ). Nectaries can be formed from the flower cover, the stamens, the ovary, the flower axis, but also outside the actual flower (extrafloral).
Position and arrangement
The petals may vary according to sheet position ( Phyllotaxis ) may be disposed whorled (cyclically) on the axis core schraubig (acyclic) or. A helical arrangement is widely regarded as a primitive feature and it is often associated with a higher number of petals. The angle between two consecutive leaves is usually close to that of the golden section of around 137.5 °.
Often only a part of the petals is screwy, while the inner petals are arranged in a whorl. These flowers are known as spirocyclic or hemicyclic . A transition from screwy to whorled arises from the fact that, as with the Yulan magnolia ( Magnolia denudata ), there is a break in leaf formation after every three leaves, so that approximately three-fold whorls are formed. A spiral sequence of leaf formation can also be seen in the development of many whorled flowers, but the leaves of a circle are so close together that a whorl is formed.
In the whorled position there is only one kind of petal in each whorl or circle. The swirling position also goes hand in hand with a reduction in the number of petals (oligomeric circles). Most whorled hermaphroditic flowers have four (tetracyclic) or five (pentacyclic) whorls: a sepal, a petal, one or two stamens and a carpel whorls. In most angiosperms, the number of leaves per whorl is also fixed; accordingly, one speaks of two-, three-, four- or five-fold flowers. If all circles have the same number of links, the flower is isomeric (as in the case of the tulip), if this is not the case, the flower is heteromeric.
The leaves of vertebrae standing on top of each other are usually on a gap (principle of alternation), that is, the limbs of the next inner whorl are in the gap between two members of the preceding whorl. With pentacyclic flowers it often occurs that the inner circle of the stamen is pushed outwards by the ovary and thus apparently becomes the outer one. This phenomenon is called obdiplostemony . All petals can be more or less fused with their own kind or with others.
If more than three planes of symmetry can be created by a flower, then it is radial symmetry (like the tulip ). If there are two planes of symmetry, it is disymmetrical , for example in Dicentra . Zygomorphic (dorsiventral) flowers have a plane of symmetry . Usually this plane of symmetry lies in the median plane of the flower (i.e. runs through the bract and stem axis of the bract), as in the mint family , less often perpendicular to the median plane, for example in the lark's spur . Asymmetrical flowers have no plane of symmetry. Strictly speaking, this also includes screw-shaped flowers, which, however, are usually viewed as radial symmetry.
The ratios of the flower parts to each other can in flowers charts graphically or in flowers formulas are presented.
Often several or many individual flowers are combined to form inflorescences .
Sexuality
The original form for the angiosperms are hermaphrodite flowers. There are also single-sex flowers (“separate sexes”), the organs of the opposite sex are usually also present as rudiments: There are staminate (“male”) and pistillate (“female”) flowers. Plants and clans with unisexual flowers can be monoecious ( monoecious , male and female flowers on one plant) or dioecious ( dioecious , male and female flowers on different plants). Polygamous plants have both hermaphroditic and unisexual flowers.
Flower induction
The flower induction is the reversal of the plant or the meristem from vegetative growth to flower formation. Triggers can be internal (endogenous) or external (exogenous) factors. The induction is irreversible, so it cannot be reversed or stopped. There are four important signaling pathways:
- Endogenous or autonomous triggers are present, but their nature is unknown. However, many species need a certain age or size before they bloom, for example many trees. In the case of the pea , the flowering time is genetically determined depending on the variety. Late flowering ecotypes of the thale cress ( Arabidopsis thaliana ) develop flowers autonomously from a certain age if there was no induction by environmental factors beforehand. The switch to flower formation occurs through activation of heterochronous genes such as FCA (Flowering Control Arabidopsis).
- For most plants, the time of flowering is determined by external factors. The most important factors are the length of the day and cold periods ( vernalization ). In terms of day length, a distinction is made between short-day plants and long-day plants, which require certain day lengths to induce flowering. The phytochrome system of the leaves plays a role as a receptor , as does the cryptochrome . The protein FKF1 also serves as a photoreceptor and measures the length of the day. These systems influence the Constans gene (CO), the expression of which alone is sufficient to trigger flower induction. A flower hormone Florigen was postulated for a long time as a signal from the leaves to the flowers, but its identity remained unknown for a long time. The FT gene (see below) is a candidate for this because, although it is transcribed in the leaves, it works in the flower meristems.
- Monocarpic plants in particular (which only bloom once) require vernalization, a longer cold period, in order to bloom. Well-known genes that are activated in cold weather are Frigida (FRI) in Arabidopsis and VRN1 and VRN2 in winter wheat. Their gene products inhibit the FLC gene, which in turn suppresses flower formation. In sum, FRI or VRN enable flower formation, even if not triggered. Other factors are usually necessary for triggering.
- Certain plant hormones , such as gibberellins , are important in stimulating flowering in the absence of long-day conditions, but the mechanism of action is still unknown.
Flower formation
After a meristem has been converted from a vegetative to a generative meristem by the heterochronous genes, the actual flower formation takes place through a large number of interacting regulatory genes.
Symmetry genes
So-called symmetry or cadastral genes determine the structure of the flower and its planes of symmetry, as well as the number of flowers and flower organs. Mutations of these genes change the architecture of the flower, the genes are often named after the phenotypes changed by such mutations : Mutations of the genes Cycloidea (CYC) or dichotoma (DICH) in snapdragons produce radial symmetry flowers instead of the usual zygomorphic ones. These mutations also occur naturally and are called pelorie . The mutation of the Cauliflower (CAL) gene in Arabidopsis and broccoli creates many branches in the inflorescence without functioning flowers. CAL inhibits the TFL1 gene, which promotes branching in the inflorescence and is a shoot identity gene. Accordingly, tfl1 mutants produce little to single-flowered inflorescences in Arabidopsis .
Flower identity genes

Within the resulting, branching inflorescence meristems are to floral meristems, if the genes Leafy (LFY) and APETALA1 (AP1) expressed are. Both code for transcription factors and belong to a group of genes that integrate the flower signals. The functioning of this and most of the other genes was obtained from Arabidopsis , snapdragons and petunia hybrids . The most important integration genes are:
- FLC (Flowering Locus C) integrates the vernalization signals and the autonomous signals. FLC is a transcription factor, its elimination leads to early flowering. The amount of FLC protein / mRNA correlates with the time of flowering. FLC represses the bloom activator SOC1.
- SOC1 (suppressor of overexpression of constans) is activated by gibberellins and via CO by Langtag, and repressed by FLC. Removal of repression by FLC is not sufficient for activation of SOC1: activation by CO or gibberellins must take place.
- Like SOC1, LFY is activated by gibberellins and by Langtag. The latter happens via SOC1 or via Agamous-Like24 (AGL24).
- FT (Flowering Locus T) is activated by long days via CO and suppressed by FLC. FT in turn activates AP1.
The last two genes in the sequence are LFY and AP1, the so-called flower meristem identity genes, but they interact with each other: AP1 is activated by LFY; however, a bloom is formed even in the absence of LFY, but not with the correct organs. The already mentioned TFL1 on the one hand and LFY and AP1 on the other hand repress each other. TFL1 serves to prevent premature flowering.
Establishment of the flower organs

The flower organs are determined according to the ABC model. These are three classes of genes that define the identity of the flower organs through interaction (organ identity genes):
- A leads to sepals
- A and B lead to petals
- B and C lead to stamens
- C leads to carpels
A and C suppress each other so that there is no overlap. In the absence of one, the other class is formed throughout the flower. The ABC model was set up in 1991 by E. Coen and E. Meyerowitz , and has since been confirmed and expanded many times.
The following genes are known in Arabidopsis :
- Class A: AP1 and AP2
- Class B: AP3 and Pistillata (PI)
- Class C: Agamous (AG)
These genes are constantly expressed in the organs that define them, not just at the beginning. AP1 is not only a flower meristem gene, but also an organ identity gene. In the early phase of flower formation, it is formed throughout the meristem, but later only in the circles of the flower envelope. Mutations lead to a change in organ identity. If A fails, carpels develop instead of sepals and stamens instead of petals (since C is now formed). Triple mutants only form normal leaves. C mutants have no limited growth; the meristem continues to grow indefinitely.
In addition, a fourth class was discovered, E, which in Arabidopsis consists of the four genes Sepallata 1 to 4 (SEP). They are expressed in the petals, stamens and carpels and are necessary for organ identity. Triple mutants (sep1 to 3) only form sepals, quadruple mutants only form leaves.
A fifth class, D, consists of genes that are responsible for the identity of the ovules . In Petunia these are the genes Floral Binding Protein (FBP) 7 and 11. Double mutants form carpel-like structures instead of ovules. In Arabidopsis , the corresponding genes are Seedstick (STK), Shatterproof1 (SHP1) and SHP2.
The majority of the flower organ genes belong to the family of the MADS box genes. MADS box genes are widely distributed in plants. They can be found, among other things, in plant homeotic genes (such as AGAMOUS and DEFICIENS), which are involved in the development of plant organism. An example of this is the definition of the flower organs. The ABCE genes are necessary, but also sufficient for the development of flower organs in vegetative organs. This could be shown with the help of ectopic expression of the genes.
How the flower identity genes, which are expressed in the entire flower meristem, control the organ identity genes has only just begun to be elucidated. First, the class B gene AP3 appears to be expressed through the combination of LFY, AP1 and the Unusual Flower Organs (UFO) gene. LFY activates the expression of AG, the C-class gene, with another coactivator, Wuschel (WUS). After activation, AG represses WUS again. This limits the growth of the flower axis.
The quartet model from Theißen 2001 brought a further step in understanding. It says that the flower organization proteins work in tetramers (hence quartet). For each organ there is therefore at least one specific tetramer. At least in yeast it has already been shown that the proteins actually form tetramers. The model already serves as a model basis for many works.
Diversity of the ABC model

Comparative studies on a number of plant species have shown that the ABC model in the form described above should only apply to the eudicotyledons . There are different characteristics, especially in the basal groups of angiosperms. In pond roses ( Nuphar ) there are no sharp limits to gene expression (“fading borders”), so that there is a transition series from calyx-like tepals via coronet-like tepals to staminodes to stamens to carpels.
The tulip forms a uniform flower envelope in two circles. Class B genes are expressed here in both circles.
The flowers of the sweet grass are very strongly derived. Nevertheless, they show the ABC model, whereby the palea is marked by A and the cavernous bodies (lodiculae) by A and B, i.e. homologous to the calyx and crown.
The dock ( Rumex ) forms a uniform, but reduced and inconspicuous flower cover. Here the B class is limited to the stamens, so that both flower envelope circles are only defined by A.
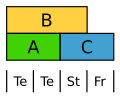
Modified ABC model of the tulip. Te Tepals
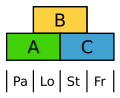
ABC model for rice. Pa palea, lo erectile tissue
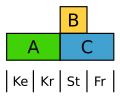
Modified ABC model for dock



In the meantime, however, the general validity of the A class as a flower organ identity genes has been questioned. As stated above, class A genes also play a role in determining the flower meristem identity. So far they could only be proven in the formation of the flower organs in Arabidopsis and they should only be of importance in close relatives. In Antirrhinum , the BC genes are sufficient for flower formation. To what extent this applies to other angiosperm groups is open.
Anthesis
The process of flowering is called anthesis, the developmental phase of the flower organs from the end of the bud state or the beginning of bud development to the beginning of withering.
Flower opening
Before opening, the flower forms a bud similar to wintering buds. The sepals serve as protection. The flower opening is one of the movements of plants . The opening of the flower bud can take place through various mechanisms:
- Reversible accumulation of ions. So far, this has only been clearly demonstrated in the case of Koch's gentian ( Gentiana acaulis ).
- Programmed cell death in defined areas of the petals.
- Water loss during the day and replenishment during the night. This occurs, for example, with Silene saxifraga , which is nocturnal and whose petals roll up during the day due to water loss.
- Different growth of the outside and inside of the petals. In the case of the tulip , the inside of the petals has a growth optimum that is 10 ° C lower than the outside, which leads to opening in the morning and closing in the evening.
An important factor for the spreading of the petals is the increase in the turgor , which is usually caused by an increase in the sugar concentration as a result of the breakdown of high-molecular carbohydrates ( starch in roses , fructans in daylilies ). This is accompanied by an expansion of the cell wall. The information on the role of plant hormones is contradictory in the literature.
The opening of the flowers is triggered by external factors. In night-blooming species, the increased humidity in the evening is often the trigger. Early spring bloomers in particular react to a rise in temperature , for example the snowdrop ( Galanthus nivalis ) or crocuses ( Crocus spp.). A third group reacts to light, like the daisy ( Bellis perennis ). The opening and closing of long-lived flowers by day or night occurs through the same mechanisms and is subject to an endogenous rhythm. The cell physiological and especially the genetic background of the opening and closing of the flowers is hardly known up to now.
The closing of the flowers can occur through differential growth or through reversible turgor changes. In these cases, repeated opening and closing is possible. Loss of turgor due to senescence leads to permanent closure of the flower.
pollination

Pollination is the transfer of male pollen to the female organs of conception: the micropyle in gymnosperms, the scars in angiosperms. Pollination is therefore not identical with fertilization. Pollination can take place with the pollen of the same individual ( self-pollination , autogamy) or with the pollen of another individual (cross- pollination , allogamy).
However, self-pollination reduces genetic variability. There are several adaptations in plants to avoid self-pollination or self-fertilization:
- Herogamy is the spatial separation of anthers and stigmas so that self-pollination is not possible. In the caper bush ( Capparis spinosa ), the entire pistil is lifted up by means of a gynophor, so that the stigma is above the anthers in the approach path of the pollinating insects.
- Dichogamy is the temporal separation of the maturity of the stamens or ovaries. Accordingly, there are pre-male ( proterandry ) and pre-female flowers ( proterogyny ). Simultaneous maturity is called homogamy.
Dichogamy and herogamy can prevent pollination within a flower, but not from one flower to a second of the same plant (geitonogamy). Therefore, many species have developed further avoidance mechanisms:


- Self incompatibility : In this case, genetic factors prevent self-fertilization. Such incompatibility systems are often also morphologically recognizable (heteromorphism): A well-known example is the heterostyly of primroses ( primula ).
There are three major adaptation syndromes, depending on the type of pollinator: pollination by wind (anemophilia), water (hydrophilicity), and animals (zoophilia). The gymnosperms are primary wind pollinators, while the first angiosperms were probably primarily animal pollinators. Wind and water pollination only developed secondarily within the angiosperms. The main features of each syndrome are:
- Anemophilia : inconspicuous flowers; reduced number of organs; Monocial or diocial often; dense, often pendulous inflorescences; little or no pollen putty; smooth pollen surface; Large surface area scars; one or a few ovules per flower; no nectar.
- Hydrophilicity : inconspicuous flowers; Monocial or diocial often; Appearance of air tissues; unwettable pollen walls; thread-like pollen grains; Large surface area scars; one or a few ovules per flower.
- Zoophilia : In the case of pollinated plants, the focus is not on the flower as a morphological unit, but on the flower as a functional unit. The blossom often corresponds to a flower (tulip), but often many blossoms are combined into one flower, which is then called pseudanthium . Examples are all the daisy family (like the daisy ) and the umbelliferae ( carrot ). It is less common that one flower forms several flowers ( meranthium ), as in the case of the iris . The main characteristics of zoophilic flowers are: hermaphrodite flowers or pseudanthia; Angiospermia; flashy color; strong fragrance; Pollen or nectar as a food supply, or deception devices; heavily carved pollen surface and lots of pollen cement.
fertilization
In the naked samers, the pollen grains get onto the micropyle of the ovules. Usually they are drawn into the pollen chamber by the drying of the pollination drop. In the pollen chamber, depending on the clan, the spermatozoids are released or the pollen tubes germinate. Up to six months can pass between pollination and fertilization (some cycads).
In angiosperms, the pollen grains get onto the stigma of the pistil. The pollen tube grows through the stylus in a special pollen tube and thus reaches the stigma to the ovules in the ovary. If there is only one uniform tissue in the style, so that pollen tubes can get from the stigma of one carpel to the ovules of another carpel, the entirety of the conductive tissue is called compitum .
Once the pollen tubes or spermatozoids have reached the egg cells, actual fertilization occurs. In angiosperms, gnetum and ephedra there is a double fertilization: in angiosperms one of the two sperm nuclei fuses with the egg cell and forms the zygote . The second fuses with the already diploid embryo sac core to form the triploid endosperm core, from which the nutritive tissue ( endosperm ) of the seeds arises. In the case of ephedra, the second sperm nucleus fuses with the abdominal canal cell of the archegonium; in the case of gnetum, the two sperm cells fuse with two gametophyte cells. Usually only one of the two zygotes develops.
After fertilization, the zygote develops into an embryo, the ovule into a seed and the flower into a fruit.
Senescence
As sexual organs in plants, the flowers are constantly being re-formed, unlike those in animals. The lifespan is precisely measured because flowers consume large resources and the stigma is also an essential entry point for pathogens. Flowers that have already been pollinated would also unnecessarily compete for pollinators with those that were not pollinated.
Pollination with pollen is an important trigger for senescence (aging) . In most species this shortens the life of the flower considerably. In many plants, pollination triggers the formation of the plant hormone ethylene , which in turn triggers the senescence of the petals. However, other species are insensitive to ethylene and their mechanism of senescence is unknown. At the organ level, the crown, stamens and stylus die off after pollination, while the ovary develops into a fruit. The death goes hand in hand with a remobilization of the ingredients, similar to the senescence of the leaves.
evolution
The evolution of the angiosperm bloom is not clear, as there are hardly any fossil precursors of the "modern" angiosperm bloom. The relationships between the angiosperms and the other seed plants are also still unclear. There are two different hypotheses about the origin of the flowers. The reason for the formation of the hermaphrodite angiosperm bloom is assumed to be the adaptation to pollination by insects (beetles); the displacement of the ovules into closed carpels would thus protect against the beetles' biting tools.
The Euanthienheorie of Arber and Parkin (1907) assumes that the ancestors of the angiosperms already had hermaphroditic flowers and therefore the angiosperm flower is a uniaxial system with lateral micro- and megasporophylls. The stamens and carpels are therefore homologous to the leaves. This theory is based on analyzes of the fossil genus Caytonia , which is believed to be the possible ancestor of angiosperms. Caytonia had pinnate micro- and megasporophylls that might have hermaphroditic flowers. In the megasporophyll, cupules, each with several ovules, sat on the side of a rachis . The microsporophylls were also pinnate, each pinna carried several groups of overgrown pollen sacs. The carpel of the angiosperms could have been created by the rachis becoming flat, the stamen by reduction to a stalk with two synangia, each with two pollen sacs.
Richard Wettstein's theory of pseudanthia assumes that the angiosperm flower emerged from an inflorescence of unisexual flowers. Thus the stamens and carpels would be side shoots and not leaves. This theory is based on the assumption that the angiosperms are descended from the Gnetopsida. The carpel was created from the bract of the flower, the second integument of the ovule from a bract below the flower. This theory is hardly supported by molecular and morphological analyzes.
Although most authors now tend to the euanthia theory, the origin of the stamen, carpel and the second integument of the ovules is still unclear.
Theories based on molecular biology
The oldest angiosperm flowers were hermaphroditic and had an undifferentiated perianth in a spiral position or more than two whorls.
Outside of the seed plants, there are no flower organ identity genes or orthologues thereof. In gymnosperms, which only form unisexual flowers, there are class B and C orthologues. Their expression is similar to that in angiosperms: C is formed in all reproductive organs, B in the male flowers. Based on these findings, several hypotheses about the origin of the hermaphroditic angiosperm bloom were made:
- According to the “out of male” hypothesis by Theissen et al. In 2002, the male cones formed female organs through a reduction in B-class expression in the upper cone area. According to the “out of female” hypothesis, the male organs formed at the lower end of the female cones. According to these two models, an inflorescence was only created after the hermaphrodite.
- The “mostly male” hypothesis is also based on male flowers, first proposed by Frohlich and Parker 2000. They discovered that the Leafy gene occurs in two copies in gymnosperms (paralogues). Leafy specifies male, needly female flowers. Needly occurs in all gymnosperms except Gnetum , but not in angiosperms. The theory now says that by deactivating the Needly gene, the male cones became hermaphroditic.
The evolution of the angiosperm bloom took place in four key events:
- Evolution of the hermaphrodite flower axis
- Evolution of the compressed flower axis and the limitation of growth: This was done by C-class genes that suppress Wuschel, the meristem maintenance gene.
- Evolution of a petaloid flower envelope
- Evolution of the classic two-circle, two-part perianth of the eudicotyledons from calyx and crown.
Soltis et al. (2007) discuss the hypothesis that at the base of the angiosperms there was not the ABC model as in Arabidopsis , but a system with fuzzy borders as in the pond rose. From this basic model, the ABC model as well as the modified forms such as the tulip can then simply have emerged through various sharpening of the boundaries.
Use by humans

In contrast to the fruits emerging from the flowers, flowers play a subordinate role as food plants for humans. The inflorescences of cauliflower and artichoke serve as vegetables, those of Cannabis sativa subsp. indica as an intoxicant. Some flowers or inflorescences are used as a salad or its decoration ( violets , daisies , nasturtiums ). Flowers, buds, flower parts or inflorescences provide spices: Mugwort ( Artemisia vulgaris subsp. Vulgaris ), lavender ( Lavandula angustifolia ), clove ( Syzygium aromaticum ), saffron ( Crocus sativus ) and hops ( Humulus lupulus ).
However, flowers play a much larger role as ornaments in the form of ornamental plants and cut flowers . As the world's largest importer of cut flowers, Germany imported goods with a wholesale value of 1.1 billion euros in 2004. The world market for cut flowers and potted plants in 2000 was 6.8 billion euros.
The goddess of the dawn ( Aurora / Eos ) scatters flowers as a symbol for the rays of light of the new day. Flowers are also the attribute of the goddess Flora . In the case of the four seasons the flowers are assigned to spring , in the case of the five senses the sense of smell . Flowers have been a sign of the transience of beauty and life since ancient times. In the still lifes, a withered blossom or fallen petals always appear as a vanitas motif. Allegories of logic and hope are sometimes depicted with flowers in their arms, as a symbol that a fruit will soon develop from the flower. Individual species have their own symbolic meanings, which can change over the centuries. The rose was and is the attribute of Venus , Dionysus and Mary , the symbol for earthly and divine love , for virgins and prostitution .
In the fine arts , blossoms and flowers as floral motifs are a frequently presented theme, primarily to express freshness and joy.
Footnotes
- ↑ “a flower as a determinate, compressed, bisexual reproductive axis composed of megasporangia (carpels), microsporangia (stamens) and a sterile perianth composed of at least one sterile laminar organ”. G. Theißen, R. Melzer: Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower . In: Annals of Botany. Volume 100, 2007, pp. 603-619.
- ↑ very similar to the definition based on a literature review in: Richard M. Bateman, Jason Hilton, Paula J. Rudall: Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers . In: Journal of Experimental Botany. Volume 57, 2006, pp. 3471-3503, doi : 10.1093 / jxb / erl128 .
- ↑ see review by Wouter G. van Doorn, Uulke van Meeteren: Flower opening and closure: a review. 2003.
- ↑ The section is based on: Wouter G. van Doorn, Uulke van Meeteren: Flower opening and closure: a review. 2003.
swell
The article is mainly based on the following sources:
- Peter Leins, Claudia Erbar: Blossom and Fruit. Aspects of morphology, evolutionary history, phylogeny, function and ecology . Schweizerbart, Stuttgart 2000, ISBN 3-510-65194-4 .
- Peter Sitte , Elmar Weiler , Joachim W. Kadereit , Andreas Bresinsky , Christian Körner : Textbook of botany for universities . Founded by Eduard Strasburger . 35th edition. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X .
The flower induction and flower formation sections are based on:
- Peter Schopfer, Axel Brennicke: Plant Physiology . Founded by Hans Mohr . 6th edition. Elsevier, Spektrum, Munich / Heidelberg 2006, ISBN 3-8274-1561-6 , pp. 501-523 ( limited preview in Google Book search).
- Thomas Jack: Molecular and Genetic Mechanisms of Floral Control . In: The Plant Cell. Volume 16, Supplement 1, 2004, pp. S1-S17, doi : 10.1105 / tpc.017038 .
- Paul K. Boss, Ruth M. Bastow, Joshua S. Mylne, Caroline Dean: Multiple Pathways in the Decision to Flower: Enabling, Promoting, and Resetting. In: The Plant Cell. Volume 16, Supplement 1, 2004, pp. S18-S31, doi : 10.1105 / tpc.015958 .
Other important literature:
- Günter Theißen, Rainer Melzer: Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower. In: Annals of Botany. Volume 100, No. 3, 2007, pp. 603-619, doi : 10.1093 / aob / mcm143 .
- Wouter G. van Doorn, Uulke van Meeteren: Flower opening and closure: a review. In: Journal of Experimental Botany. Volume 54, No. 389, 2003, pp. 1801-1812, doi : 10.1093 / jxb / erg213 (section flower opening).
Individual evidence
- ^ Friedrich Kluge , Alfred Götze : Etymological dictionary of the German language . 20th edition. Edited by Walther Mitzka . De Gruyter, Berlin / New York 1967; Reprint (“21st unchanged edition”) ibid 1975, ISBN 3-11-005709-3 , p. 87.
- ↑ a b Gerhard Wagenitz : Dictionary of Botany. The terms in their historical context . 2nd expanded edition. Spectrum Academic Publishing House, Heidelberg / Berlin 2003, ISBN 3-8274-1398-2 , p. 47 f .
- ↑ Peter Schütt , Hans Joachim Schuck, Bernd Stimm (ed.): Lexicon of tree and shrub species. The standard work of forest botany. Morphology, pathology, ecology and systematics of important tree and shrub species . Nikol, Hamburg 2002, ISBN 3-933203-53-8 , pp. 83 (reprinted 1992).
- ^ Manfred A. Fischer, Wolfgang Adler, Karl Oswald: Excursion flora for Austria, Liechtenstein and South Tyrol . 2nd, improved and enlarged edition. Province of Upper Austria, Biology Center of the Upper Austrian State Museums, Linz 2005, ISBN 3-85474-140-5 , p. 90 .
- ^ Rudolf Schubert , Klaus Werner, Hermann Meusel (eds.): Exkursionsflora for the areas of the GDR and the FRG . Founded by Werner Rothmaler. 13th edition. tape 2 : vascular plants . People and knowledge, Berlin 1987, ISBN 3-06-012539-2 , pp. 24 .
- ^ Siegmund Seybold (Ed.): Schmeil-Fitschen interactive . CD-ROM, version 1.1. Quelle & Meyer, Wiebelsheim 2002, ISBN 3-494-01327-6 .
- ^ Peter Sitte , Elmar Weiler , Joachim W. Kadereit , Andreas Bresinsky , Christian Körner : Textbook of botany for universities . Founded by Eduard Strasburger . 35th edition. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X , p. 750 .
- ↑ Herder Lexicon of Biology . CD-ROM. Spektrum Akademischer Verlag, Heidelberg 2003, ISBN 3-8274-0354-5 , keyword “Blossom”.
- ^ Peter Sitte , Elmar Weiler , Joachim W. Kadereit , Andreas Bresinsky , Christian Körner : Textbook of botany for universities . Founded by Eduard Strasburger . 35th edition. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X , p. 720, 723 .
- ^ Katja E. Jaeger, Alexander Graf, Philip A. Wigge: The control of flowering in time and space. In: Journal of Experimental Botany. Volume 57, No. 13, 2006, pp. 3415-3418, doi : 10.1093 / jxb / erl159 .
- ↑ Laurent Corbesier, George Coupland: The quest for-pile: a review of recent progress. In: Journal of Experimental Botany. Volume 57, No. 13, 2006, pp. 3395-3403, doi : 10.1093 / jxb / erl095 .
- ^ Enrico S. Coen, Elliott M. Meyerowitz: The war of the whorls: genetic interactions controlling flower development. In: Nature. Volume 353, 1991, pp. 31-37, DOI: 10.1038 / 353031a0 .
- ↑ a b c d e f g h i Günter Theißen, Rainer Melzer: Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower. In: Annals of Botany. Volume 100, No. 3, 2007, pp. 603-619, doi : 10.1093 / aob / mcm143 .
- ^ A b Douglas E. Soltis, André S. Chanderbali, Sangtae Kim, Matyas Buzgo, Pamela S. Soltis: The ABC Model and its Applicability to Basal Angiosperms . In: Annals of Botany. Volume 100, No. 2, 2007, pp. 155-163, doi : 10.1093 / aob / mcm117 .
- ↑ Video of blossoms opening on YouTube .
- ↑ Hilary J. Rogers: Programmed Cell Death in Floral Organs: How and Why do Flowers Die? In: Annals of Botany. Volume 97, No. 3, 2006, pp. 309-315, doi : 10.1093 / aob / mcj051 .
- ^ EA Newell Arber, John Parkin: On the origin of the angiosperms. In: Journal of the Linnean Society of London, Botany. Volume 38, No. 1, 1907, pp. 29-80, DOI: 10.1111 / j.1095-8339.1907.tb01074.x ( digital copy ).
- ^ Richard Wettstein: Handbook of Systematic Botany. Volume 2, F. Deuticke, Vienna 1907.
- ↑ Michael W. Frohlich, David S. Parker: The mostly male theory of flower evolutionary origins: from genes to fossils. In: Systematic Botany. Volume 25, No. 2, 2000, pp. 155-170, DOI: 10.2307 / 2666635 ( PDF file ).
- ↑ a b Richard M. Bateman, Jason Hilton, Paula J. Rudall: Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers. In: Journal of Experimental Botany. Volume 57, No. 13, 2006, pp. 3471-3503, doi : 10.1093 / jxb / erl128 .
- ^ Wolfgang Franke : Nutzpflanzenkunde. Usable crops of the temperate latitudes, subtropics and tropics . 4th revised and expanded edition. Thieme, Stuttgart / New York 1989, ISBN 3-13-530404-3 .
- ↑ florist.de ( Memento from November 5, 2007 in the Internet Archive )
- ↑ ianywhere.com, with mobile technology successfully in the flower market , accessed on 12 December 2007
- ↑ Lucia Impelluso: Nature and its symbols. Plants, animals and mythical creatures . Parthas Verlag, Berlin 2005, ISBN 3-936324-03-4 .
- ^ Marianne Beuchert : Symbolism of the plants . Insel Verlag, Frankfurt / Leipzig 2004, ISBN 3-458-34694-5 , p. 279 .
further reading
- Dieter Hess: The blossom. An introduction to the structure and function, ecology and evolution of flowers, with instructions for simple experiments . 2nd improved and supplemented edition. Eugen Ulmer, Stuttgart (Hohenheim) 1990, ISBN 3-8001-6434-5 (generally understandable introduction also for the layman, out of print).
- Special Issue: Major Themes in Flowering Research. In: Journal of Experimental Botany. Volume 57, No. 13, 2006 ( online ).
- Douglas E. Soltis, James H. Leebens-Mack, Pamela S. Soltis (eds.): Developmental Genetics of the Flower (= Advances in Botanical Research . Volume 44 ). Academic Press, San Diego et al. a. 2006, ISBN 0-12-005944-4 (English, online ).