
Rose plants
| Rose plants | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Dog rose ( Rosa canina ) |
||||||||||||
| Systematics | ||||||||||||
|
||||||||||||
| Scientific name | ||||||||||||
| Rosaceae | ||||||||||||
| Yuss. |
The rose plants (Rosaceae) are a plant family of the kernel eudicotyledons . The family is distributed worldwide, with a focus on the northern hemisphere . The representatives are herbaceous plants, shrubs or trees and usually have showy, hermaphrodite flowers with a double flower envelope and a clearly pronounced flower cup . In addition to the name-giving roses ( pink ) used as ornamental plants, the family also includes many well-known types of fruit such as apples , pears , blackberries , strawberries and raspberries as well as stone fruits with cherries , plums , plums , almonds and others.
description
Vegetative characteristics
The representatives of the rose family are trees , shrubs or herbaceous plants , whereby the shrubby growth form is regarded as the original within the family. Large trees with heights of 25 to 30 meters are rare and only occur in a few genera (such as Eriobotrya , Sorbus , Prunus ). Woody plants can be evergreen or deciduous. Some clans have shoot thorns . Spines are common in some genera , especially Rosa and Rubus . Herbaceous representatives are mostly persistent and form underground vertical rhizomes or horizontal rhizomes as Überdauerungsorgane.
The leaves are mostly alternate. In the case of the leaf shape , the simple leaf is considered to be the original form in the family, compound leaves are considered to be derived. Compound leaves are usually paired or unpaired pinnate leaves and come in around 30 genera. Opposite leaves occur only in the genera Coleogyne , Rhodotypos and Lyonothamnus . The presence of stipules is considered an original feature. In the Spiraeoideae , the stipules were lost independently of one another in several lines of development. The stipules are often fused to the base of the petiole . There are usually two glands at the apical end of the petiole. The leaf margin is often serrated, rarely entire. In some genera, water gaps or guttation were observed. In wood , the basic fabric consists mainly of tracheid fibers , more rarely also of libriform fibers .
Inflorescences and flowers


The flowers are mainly in racemose or panicle inflorescences , spikes , heads , solitary flowers are rare.
The flowers are radial symmetry and mostly hermaphroditic , rarely unisexual due to reduction. A flower cup (hypanthium) is always present and differently pronounced. As a rule, there is a double flower envelope , the calyx and crown are usually five-fold. The sepals sit on the edge of the flower cup and are rather small. Sometimes an outer cup is also formed. The petals are usually conspicuous, the predominant color is white, in some groups also yellow. The lack of petals, such as in the genus Sanguisorba, is a derived characteristic and mostly occurs in wind-pollinated groups.
In the case of the stamens , a number of over 10 to many is considered to be original ( secondary polyandry ), there are also fewer secondary. Often there are around 20. They are always free and not grown together. They are usually separated from the carpels by a nectar- secreting disc. The pollen grains are two-celled when released and emerge individually as monads. They are more spherical, generally tricolporate (have three germinal furrows / pores) and with long furrows.
On the carpels , the number from one to five is considered original, there can also be many. The carpels are free (apocarp) or fused together. They sit on the flower base (such as strawberries ) or they are surrounded by the recessed flower cup (such as roses ) or grown together with this (such as apples ). There are all transitions from the upper to the lower ovary . There are usually two anatropic ovules per carpel , which have one or two integuments , more rarely - in original clans - several ovules. The embryo sac develops according to the Polygonum type, the embryo according to the Asterad type. The styles are free with the exception of some representatives of the Pyreae tribe .
The flower formula that applies to most species is thus:

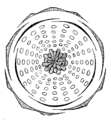
Floral diagram of Rosa tomentosa

Flower diagram of Prunus padus

Flower diagram of Sanguisorba officinalis

Flower diagram of Spiraea hypericifolia




Fruits and seeds


The fruits of the rose family are very diverse. In the past, the fruits and the associated flower morphology were used as the main characteristic for the systematic classification of the family, while more recent molecular genetic studies undertake group formation based on the genetic similarities between the species.
Accordingly, the four subfamilies Spiraeoideae, Rosoideae, Maloideae and Prunoideae can be found in the identification literature, while new specialist literature on genetics or the secondary metabolism of Rosaceae uses the updated classifications in parallel to this classic system.
According to the classical view, the Rosaceae can be divided into the following four subfamilies or morphological groups based on their fruits:
- Spiraeoideae , the typical fruit of which is the follicle . The tree species include, for example, the originally considered species Aruncus dioicus (goat's beard) and Agrimonia eupatoria (ormennig).
-
Rosoideae (rose-like), the carpels of which, as in Filipendula ulmaria (meadowsweet), form individual nut fruits or various collective fruits as in the strawberry Fragaria .
- The dry fruits of the Rosoideae often have an elongated stylus with bristles or barbs, which, like the brook avens ( Geum rivale ), serve to spread the fruit through animals ( zoochory ).
- In juicy, fleshy fruits such as strawberries, the numerous carpels often sit individually ( apocarp ) on the clumpy, bulging flower base and also form nutlets. During the ripening process, the flower base expands like a balloon and turns signal red due to secondary dyes. The strawberry is a dummy fruit because its edible tissue is not formed from the carpel.
- The pulp of the rose hip ( Rosa canina ) is also formed from the flower base. It encloses many small nuts that represent the actual fruit.
- Maloideae (apple-like), which are based on the phylogenetically more original collecting bellows. The apple fruit is the Gynoeceum not upper constant ( epigyn but) sunk into the floral axis, where it forms from the five carpels the so-called. Cored, the everyday language as "slugs" is called. When the fruit ripens, this gynoeceum is overgrown by the surrounding axial tissue, so that in the apple fruit it is not the carpel that is consumed, but parenchymatic stem axis tissue. The apple fruit is therefore one of the false fruits .
- Prunoideae (stone fruit plants) form a seed that is surrounded by the inner, lignified layer of the pericarp (core or stone), while the outer area of the fruit swells and is accentuated in color by the storage of secondary substances such as anthocyanins. A hypanthium is typical ofthe stone fruit family, in which the gynoeceum sits in the cup-shaped retracted flower base (receptaculum). Such a position of the gynoeceum is called perigyn .
The seeds often have a solid, sometimes thick-walled seed coat . It is formed from the so-called integuments of the ovule. The seed coat protects the embryo, which mainly consists of the two flat to thick cotyledons . In the ripe seeds of the Rosaceae, an additional nutrient tissue ( endosperm ) is only sometimes preserved or only a thin layer is left.
Chromosome numbers
The basic chromosome numbers of Rosaceae are x = 7, 8, 9, 15, and 17. Originally 7 or 9 are considered. The base number 8 developed once in the Rosoideae and several times in the Spiraeoideae, the base number 17 developed once in the Pyreae.


ingredients
The Rosaceae are rich in tannins that are diffusely distributed in the tissue or occur in tannin idioblasts . They are predominantly condensed tannins (pro anthocyanidins ), in the subfamily Rosoideae also gallo- and ellagitannins occur to a greater extent . The monomeric precursors of the tannins such as catechins and flavonol glycosides of kaempferol and quercetin also occur. Ellagic acid occurs only in the Rosoideae.
Pentacyclic triterpenes such as oleanolic acid and ursolic acid are widespread in the cuticular waxes . Triterpene alcohols (such as lupeol , betulin ) are often found in the bark . Triterpenic acids, which are esterified with sugars via the carboxy group , are referred to as pseudosaponins . One example is Tormentol. Real saponins are also found. The seeds are usually free of starch and store proteins and fatty oil.
Cyanogenic glycosides such as amygdalin or prunasin are widespread within the family .
Sorbitol is stored in the leaves and fruits of many representatives as a carbohydrate reserve, but is absent in many representatives of the Rosoideae. Sorbitol is also often the carbohydrate transported in the phloem .
The walls of the seed-shell epidermis of the pome fruit plants are often heavily mucous, especially in the quince ( Cydonia oblonga ). Fruit acids are sometimes found in high concentrations in fruits, especially malic and citric acid , and sometimes succinic acid , as well as ascorbic acid (vitamin C).
ecology
pollination
The majority of the rose family has disc and shell flowers , in which the individual flower is relatively large and acts as a pollination unit. Brush flowers are seldom represented, here the individual flowers are relatively small and stand together in head-like or spike-like inflorescences. This type of flower occurs mainly in the Sanguisorbeae. Flies and short-tongued bees are the main pollinators . Long-tongued bees, beetles and butterflies are of less importance than pollinators . Some genera of the Sanguisorbeae are wind pollinated .
Cross-pollination is achieved in many species by a more or less pronounced pre-femininity ( protogyny ), most sexually reproducing species are also incompatible .
Polyploidy and Reproductive Ecology
Polyploidy is found in around a third of the genera . Polyploidy often occurs in the species-rich genera. Examples are Alchemilla (1000 species, up to 28-ploid), Potentilla (up to 16-ploid), Rosa and Rubus .
Apomixis has developed in some genera , it is mostly coupled with polyploidy. The most common form of apomixis is apospore , in which the embryo sac develops from an unreduced vegetative cell. Diplosporia also rarely occurs , in which the embryo sac develops from an unreduced generative cell derived from the embryo sac mother cell . For the seed set, however, pollination and fertilization of the endosperm are usually necessary. Therefore it is in principle also possible for the unreduced egg cell to be fertilized and for an increase in the number of chromosomes to take place.
In contrast to the other members of the family, a large part of the polyploid rose family is self-compatible, i.e. a plant can pollinate itself.
Hybridization occurs frequently in the rose family, and in the Pyreae even across genus boundaries. The apomixis often associated with hybrid formation led to the formation of diploid-triploid-tetraploid networks of species in some genera. Hybrids became fertile through polyploidization, or triploid clans can reproduce apomictically.
Propagation ecology
In the rose family there are four main types of spreading of the diaspores , which are closely related to the fruit types: meaty fruits are eaten by animals, the seeds excreted again (endozoochory). In the case of trees, these are mostly birds, bats or other mammals, in the case of strawberries it is likely to be snails, in the case of aremonia , due to the presence of elaiosomes, ant spread ( myrmecochory ) is suspected. Epizoochoria occurs in species that have hooked on hypanthium or style. There are also seed ejectors and, rarely, anemochories (such as Dryas and Geum ).
Symbioses
Vesicular-arbuscular mycorrhiza occurs in the rose family about as often as in all Rosopsida , while ectomycorrhiza was only found in a few woody species of the Dryadoideae, Pyreae, Prunus and Rosa . The Dryadoideae enter into a symbiosis with the nitrogen-fixing bacteria of the genus Frankia .
Distribution and locations

The representatives of the Rosaceae are distributed worldwide. Only three of the genera are cosmopolitan , namely Prunus, Alchemilla and Rubus , although only with Prunus is it certain that it occurs naturally on all continents without human influence. 16 genera are distributed over the entire temperate zone of the northern hemisphere, most of which also extend into the Mediterranean or even tropical climates, here mostly in the montane altitude zone further south. Geum and Agrimonia also extend to the southern hemisphere. Only two genera occur only in the southern hemisphere, Acaena and Oncostylis . Most of the endemic genera are found in Asia and 17 in North America, with only a few in South America (3), Africa (5) and Europe (1).
Rose plants grow in a variety of locations, from semi-deserts to lowland rainforests and open, alpine vegetation. A large number of species can be found on wooded mountain slopes of medium altitude and moderate latitude.
meaning

The family is home to many ornamental and useful plants. Species used as fruit are particularly: Quince ( Cydonia oblonga ), Japanese loquat ( Eriobotrya japonica ), garden strawberry ( Fragaria × ananassa ), forest strawberry ( Fragaria vesca ), cultivated apple ( Malus domestica ), medlar ( Mespilus germanicus ) , Apricot or apricot ( Prunus armeniaca ), sweet cherry ( Prunus avium ), sour cherry ( Prunus cerasus ), plum , plum and mirabelle plum (subspecies of Prunus domestica ), blackthorn ( Prunus spinosa ), pear ( Pyrus communis ), Cloudberry ( Rubus chamaemorus ), blackberries ( Rubus fruticosus agg.), Raspberry ( Rubus idaeus ) and sweet mountain ash ( Sorbus aucuparia var. Edulis ). Several species are also used for medicinal purposes. Tanning drugs predominate here: the rhizome of Potentilla erecta , leaves of blackberry, lady's mantle and ormint.
Systematics
The Rosaceae are within the order of the rose-like (Rosales) the sister group of all other families of the Rosales.
The family Rosaceae includes around 90 genera with a total of around 3000 species . In the past, fruit was the main characteristic for the systematic structuring of the family, while today attempts are made to include molecular genetic correspondences between the genera in the system. As a result of the innovation, the old subfamily names Prunoideae and Maloideae have been partially discarded in the relevant specialist literature; the assigned plants were moved to the Triben Pyrinae (pome fruit plants, Spiraeoideae ) and Prunus (stone fruit plants, Spiraeoideae). According to the newer view, the stone and pome fruit plants no longer form their own subfamilies.
To laypeople, this simultaneity of two different systems can be confusing; However, they each pursue their own objective and can therefore not be assessed as right or wrong, but only according to their practical use with regard to a task: The morphological systematics primarily serves as an orientation for the anatomy of the plants and the plant identification, while one is aware of hopes for more valid knowledge about the phylogenetic relationships of the Rosaceae from the molecular perspective.
The groups listed here follow the molecular genetic relationships and are each natural relationship groups ( monophyletic ). Nevertheless, the more precise relationships between the subfamilies and especially between the tribes of the Spiraeoideae are not yet sufficiently known.














|
The traditional division into the four sub-families Prunoideae, Pyrinae, Spiraeoideae and Rosoideae goes back to Wilhelm Olbers Focke in 1888. The division into tribes comes largely from Maximowicz for Spiraeoideae (1879), Focke for Rosoideae (1888) and Koehne for Maloideae (today: Pyrinae) (1890), the Prunoideae were never divided into tribes. A division of the von Hutchinson family (1964) into 20 tribes without subfamilies could not prevail. The revision of the family in 2007 with the incorporation of the previous subfamilies Prunoideae and Pyrinae into the Spiraeoideae, as well as the establishment of a new subfamily Dryadoideae, aimed at the formation of monophyletic taxa.
Fossil history

The Rosaceae have been well documented by numerous fossils from North America and Europe since the Eocene . From the Neogene there are also finds from Asia, the Arctic, North Africa and some Gondwana regions. Well-studied finds come from the Okanagan Highlands (Rocky Mountains) from the early to mid-Eocene. There are recent genera that were already fully developed in the Eocene, species that resemble recent genera and extinct genera that resemble higher taxa in their characteristics . Examples of the latter are Paleorosa and Stonebergia . The genus Prunus is also well represented with fossils that are already very similar to today's representatives.
The oldest in situ rosacea pollen is known from Paleorosa . The morphological features of the genus are between the classic subfamilies Pyrinae and Spiraeoideae . The fruits of Quintacava are similar to the pome fruits . Genera widespread today, which have also been documented since the Eocene, are, for example, Rubus , Crataegus and Sorbus .
In Europe, the rose plants are an important element of the flora in the Neogene. Widespread genera here are Rosa , Crataegus , Rubus , Mespilus , Sorbus and Prunus .
literature
- Daniel Potter, Torsten Eriksson, Rodger C. Evans, SH Oh, Jenny EE Smedmark, David R. Morgan, M. Kerr, Kenneth R. Robertson, Matthew P. Arsenault, Timothy A. Dickinson, Christopher S. Campbell: Phylogeny and classification of Rosaceae. In: Plant Systematics and Evolution. Volume 266, 2007, No. 1-2, pp. 5-43. doi : 10.1007 / s00606-007-0539-9 (section systematics)
- C. Kalkman: Rosaceae , in: Klaus Kubitzki (Ed.): The Families and Genera of Vascular Plants - Volume VI - Flowering Plants - Dicotyledons - Celastrales, Oxalidales, Rosales, Cornales, Ericales. Springer, Berlin 2004, p. 343ff. ISBN 978-3-540-06512-8 (Characteristics, Ecology, Distribution and Locations sections)
- Cuizhi Gu, Chaoluan Li, Lingdi Lu, Shunyuan Jiang, Crinan Alexander, Bruce Bartholomew, Anthony R. Brach, David E. Boufford, Hiroshi Ikeda, Hideaki Ohba, Kenneth R. Robertson, Steven A. Spongberg: Rosaceae. : P. 46–434 - same text online as the printed work , PDF 4.6 MB , In: Wu Zheng-yi, Peter H. Raven (Ed.): Flora of China , Volume 9 - Pittosporaceae through Connaraceae , Science Press and Missouri Botanical Garden Press, Beijing and St. Louis, 2003, ISBN 1-930723-14-8 . (Sections Characteristics and Systematics)
- Dietrich Frohne, Uwe Jensen: Systematics of the plant kingdom with special consideration of chemical characteristics and plant drugs . 4th revised edition. Gustav Fischer, Stuttgart / Jena / New York 1992, ISBN 3-437-20486-6 (section ingredients).
- ML de Vore, KB Pigg: A brief review of the fossil history of the family Rosaceae with a focus on the Eocene Okanogan Highlands of eastern Washington State, USA, and British Columbia, Canada. Plant Systematics and Evolution, Volume 266, 2007, pp. 45-57. doi : 10.1007 / s00606-007-0540-3 (section fossil history)
- Daniel Potter, Barbara Ertter: Rosaceae at Jepson eFlora , 2012, description and key. (Section systematics)
- The Rosaceae family at DELTA . (Section description)
Individual evidence
- ↑ a b D. Frohne, U. Jensen: Systematics of the plant kingdom with special consideration of chemical characteristics and plant drugs . 4th edition, G. Fischer, Stuttgart, Jena, New York 1992, p. 147ff. ISBN 3-437-20486-6
- ↑ Khidir W. Hilu, Thomas Borsch, Kai Müller, Douglas E. Soltis, Pamela S. Soltis, Vincent Savolainen, Mark W. Chase, Martyn P. Powell, Lawrence A. Alice, Rodger Evans, Hervé Sauquet, Christoph Neinhuis , Tracey AB Slotta, Jens G. Rohwer, Christopher S. Campbell, Lars W. Chatrou: Angiosperm phylogeny based on matK sequence information . In: American Journal of Botany , Volume 90, 2003, pp. 1758-1776. (on-line)
- ↑ MA Fischer: What is the natural system? Reflections on the concept of the biological system in the age of molecular physics. In: Verh. Zool.-Bot. Gesellschaft Österreich , issue no. 148/149, 2012, pp. 323–362
- ↑ D. Potter, T. Eriksson, RC Evans, S. Oh, JEE Smedmark, DR Morgan, M. Kerr, KR Robertson, M. Arsenault, TA Dickinson, CS Campbell: Phylogeny and classification of Rosaceae . In: Plant Systematics and Evolution , Volume 266, 2007, pp. 5-43. doi : 10.1007 / s00606-007-0539-9
- ↑ Daniel Potter, Barbara Ertter: Rosaceae at Jepson eFlora , 2012, description and key.
- ↑ Cuizhi Gu, Chaoluan Li, Lingdi Lu, Shunyuan Jiang, Crinan Alexander, Bruce Bartholomew, Anthony R. Brach, David E. Boufford, Hiroshi Ikeda, Hideaki Ohba, Kenneth R. Robertson, Steven A. Spongberg: Rosaceae. : P. 46–434 - same text online as the printed work , PDF 4.6 MB , In: Wu Zheng-yi, Peter H. Raven (Ed.): Flora of China , Volume 9 - Pittosporaceae through Connaraceae , Science Press and Missouri Botanical Garden Press, Beijing and St. Louis, 2003, ISBN 1-930723-14-8 .
- ^ David John Mabberley: The Plant Book. A portable dictionary of the higher plants . Cambridge University Press 1987. page 469. ISBN 0-521-34060-8
- ↑ Rosaceae in the Germplasm Resources Information Network (GRIN), USDA , ARS , National Genetic Resources Program. National Germplasm Resources Laboratory, Beltsville, Maryland. Retrieved April 11, 2017.
Web links
- The Rosaceae family on the AP website . (engl.)