
Hominization

As Hominisation (also Anthropogenesis rarely Anthropogenie ), the evolutionary referred emergence of the feature structure that the anatomically modern humans ( Homo sapiens is characteristic).
In the course of hominization, both his physical characteristics and his mental abilities have developed. These include in particular the upright gait , the set of teeth with a shortened, parabolic dental arch and small canine teeth, as well as the late onset of sexual maturity ; furthermore, the enlargement of the brain and the associated cultural and social abilities that distinguish human beings and their ancestors from other great apes.
Based on fossil found and molecular dating of the beginning of hominization is dated to the time before 7-5 million years ago, when the line of development of the chimpanzee from the leading to humans line of development Hominini separated.
Even in the present, human evolution has not come to a standstill.
Anatomical characteristics and behavior
The following features distinguish the American anatomist C. Owen Lovejoy , according to the anatomically modern humans (the Homo sapiens ) from the other species of great apes : the upright gait , a large neocortex (an area of the cerebral cortex ), reduced incisors and canines , a unique sexual and reproductive behavior and material culture. Chris Stringer also referred to a large, round skull (due to the strongly arched parietal bone ), at most very small bulges above the eyes and a protruding bone (the chin ) on the lower jaw .
Leslie Aiello named the following characteristics in 2014: “Large, straight torso, elongated legs, large brain, reduced sexual dimorphism , increased meat consumption, unique characteristics of life history (e.g. extended embryonic and fetal phases as well as longevity), further tool manufacture and increased social Cooperation. ”The skin also shows major genetic and morphological differences compared to chimpanzees.
One of the oldest human-like anatomical features is the shape of the anvil in the middle ear , which has already been documented for Australopithecus africanus and Paranthropus robustus and which probably differed from the shape of the anvil in the chimpanzees in their last common ancestor. In the 3.3 million year old fossil DIK 1-1 , a juvenile Australopithecus afarensis , the thoracic spine already consisted of 12 vertebrae , as in most later hominini, and not of 13 vertebrae, as in most chimpanzees.
A comparison of skeletal finds of early homosexuals from different regions of Africa and Georgia - without taking into account their assignment to certain species - gave indications of considerable regional differences in size and weight (see table).
| Location and age | height | body weight |
|---|---|---|
| Koobi Fora , younger than 1.7 mya | 157 - 178 cm | 60 - 81 kg |
| South African caves, 1.8 to 1.4 mya | 136-159 cm | 32 - 62 kg |
| Olduvai Gorge , 1.75 mya | 134-162 cm | 38 - 65 kg |
| Dmanissi , 1.8 mya | 147 - 153 cm | 46 - 53 kg |
| Koobi Fora, older than 1.8 mya | 148 - 168 cm | 39 - 68 kg |
Walking upright

left: Australopithecus (3.6 million years old)
middle: Homo erectus (1.5 million years old)
right: anatomically modern human being
Based on the skeletal characteristics of many fossil finds could be proved to the upright, bipedal gait ( bipedalism ) in the family of great apes evolved much earlier than the high magnification of the brain. In 1988, C. Owen Lovejoy described the transition to an upright gait as the most obvious change in anatomy that has been demonstrated in all of evolutionary biology.
Richard Leakey According to this change is so unique, "that we are entitled to all kinds of bipedal apes than human [human] to call." Such a broad definition is controversial, however, in professional circles, as not at least sometimes going all the fossils of upright apes - are to be assigned to similar individuals of the immediate ancestral row of humans and the upright gait therefore seems to have developed several times independently of each other in the form of the great apes.
This was possible because as early as in the Middle Miocene - around 10 million years ago - the ancestors of the great apes had changed from the previously arborical-quadrupede mode of locomotion (leaning forward and walking four-legged over branches) into a suspensory mode of locomotion (standing under the branches), which a gradually progressive remodeling of arms, legs and torso as a prerequisite. This redesign, which can be demonstrated in many primate species of the Miocene, is in turn considered to be a significant pre-adjustment for the later transition to a ground-living, two-legged, upright mode of locomotion. The most striking changes to the foot concerned the structure and interaction of the toe and metatarsal bones , which were originally a grasping organ.
From the bone structure of fossils such as " Little Foot " and " Lucy " it could be deduced that even individuals of the genus Australopithecus could walk upright over long distances. This interpretation was confirmed in 1979, when 3.6 million year old fossil footprints of Australopithecus afarensis were discovered in Laetoli , which had preserved its bipedal locomotion. However, it remained controversial whether Australopithecus afarensis - comparable to a chimpanzee - walked with the knee and hip joint constantly bent or, like anatomically modern humans, more energetically with a bent knee. It was not until 2010 that a biomechanical experiment provided evidence that the fossilized footprints preserved an imprint profile that largely resembles that of people living today: When walking upright, the imprint depth of the toes and heels are approximately the same; in chimpanzee-like walking, the toes press deeper into the ground than the heel. According to this, a human-like upright gait - in terms of motion sequences and energy efficiency - developed long before the genus Homo emerged ; nevertheless, individual ontogenetic adaptations of the foot as a result of frequent climbing can still be demonstrated in anatomically modern humans. Around two million years ago, in addition to the upright species, closely related species existed in South Africa, whose bone structure primarily shows an adaptation to climbing.
The oldest fossil evidence of a foot comparable in function to humans is a fully preserved, 3.2 million year old metatarsal bone of Australopithecus afarensis from Hadar (archive number AL 333-160), the characteristics of which are both the presence of a longitudinal vault and a Reveal transverse vault. As the straight, non-curved shape of the bone and the still recognizable attachment points of the muscles show, the transition from a grasping foot optimized for climbing in branches with a slightly curved metatarsal to an arch that acts as a "shock absorber" when walking upright was already well advanced.
Further direct evidence of a form of upright, two-legged locomotion, which is essentially comparable to that of people living today, is also provided by Laetoli, 1.51 to 1.53 million years old and attributed to Homo erectus . According to the analyzes, the toes were relatively short, the big toe - unlike in monkeys and still in the 4.4 million year old fossil Ardi - aligned parallel to the other toes; the feet were arched a little upwards, as is the case with people alive today . As you walked, the weight shifted from the heel to the ball of the foot . From the distances between the footprints and their size and depth, the researchers concluded that the creators of the footprints were similar in height and weight to people today.
However, it has not yet been clarified whether the ability of humans to run for endurance, which is unique among primates - for example, to run a marathon - is a mere accompaniment of the ability to walk upright or whether it developed as a result of separate selection pressure .
There are numerous hypotheses about the origin of the upright gait , which are not necessarily mutually exclusive.

.jpg)
The hypothesis of the development of upright walking on trees
According to a research team led by Susannah Thorpe from the University of Birmingham and Robin Crompton from the University of Liverpool, the upright gait could have already developed in the ancestors of humans, who still predominantly lived on trees, in order to improve the fruit on the ends of thin branches, for example to be able to achieve.
The researchers had observed orangutans in Sumatra for a year and reported on them in 2007. These great apes spend their entire lives in trees and therefore could serve as a model of how our ancestors lived millions of years ago. The analysis of around 3000 movements showed that the orangutans move on very thin twigs on two legs, holding onto branches hanging over them with their hands and balancing their weight with their arms. On the other hand, they tend to hang on medium-thick branches, and very thick branches are mastered in the quadruped walk. According to this line of argument, our ancestors would have been walking on two legs, at least at times, much earlier than previously assumed. Only later, when many African rainforests gradually disappeared during a dry period, would they have reacted by “moving” to the ground, where they further developed and finally perfected the upright gait.
The other line, which led to today's chimpanzees and gorillas , however, developed a quadruped gait on the knuckles (the so-called ankle gait ) in order to be able to get quickly from one tree to the next in the thinned forests.
This hypothesis is supported by the fact that some hominin finds come from areas that were clearly forested during their lifetime. This applies, for example, to the Orrorin discovered in 2000 and to Australopithecus finds such as " Lucy ". This hypothesis is also supported by detailed analyzes of the 4.4 million year old Ardi skeleton of Ardipithecus ramidus . Friedemann Schrenk described his mode of locomotion as follows: “The construction of Ardi's foot is exciting. It was built so that she could gracefully walk on the branches. So she didn't clamber along them like chimpanzees do. ”A comparison of the wrist bones of chimpanzees and gorillas also showed that their ankle gait developed independently of each other, so that their last common ancestor did not yet have this walking posture. From this it was deduced that the hominini also do not descend from ancestors with ankle gait. The find by Danuvius guggenmosi published in 2019 was also interpreted as confirmation of this hypothesis.
The hypothesis of the relationship between walking upright and eating
A hypothesis about the emergence of the upright gait as a result of eating in a certain posture ("postural feeding hypothesis") was brought up by the paleoanthropologist Kevin D. Hunt of Indiana University . This theory asserts that chimpanzees are regularly bipedal when eating. On the ground they would reach up to get fruit hanging from smaller trees, and on the trees the ability to temporarily bipede would be used by reaching for a branch above them. These two-legged movements developed into more frequent habits, according to the theory.
Hunt's hypothesis can be described as a forerunner to the hypothesis of the development of upright walking on trees, but it was "falsified" in 2009 by analyzing the physique of Ardipithecus ramidus , as C. Owen Lovejoy explicitly noted.
Savannah hypothesis
The savannah hypothesis was the assumption that the evolution of the bipedia started about 7 to 8 million years ago in the great apes because the species that were still living in forests at that time moved their habitat to open, treeless savannahs and walked there upright would have found; even Jean-Baptiste Lamarck (1744-1829), this had suspected. This hypothesis is now considered refuted due to numerous fossil finds, since the earliest upright genera, such as Sahelanthropus and Orrorin, which were associated with the hominini , were not pure savannah inhabitants, but lived in gallery forests . Also Ardipithecus ramidus lived in an environment "comprised habitats from closed woodland and fanned trees." The
The thermoregulatory model, which the zoologist and evolutionary biologist Peter Wheeler from Liverpool John Moores University from 1983 onwards, is therefore considered to be irrelevant for the development of upright gait . He had argued, among other things, that the bipedia developed because this posture protected the brain from overheating when it was in the open savannah: the higher the body of an ape rose above the ground, the more likely it was to be exposed to higher, cooling wind speeds been.
Nonetheless, researchers assume that walking upright in the African hominini's habitat, which was proven to be hot 4 million years ago, was also beneficial for climatic reasons, and that the thermoregulatory aspects highlighted by Wheeler in the 1980s and 1990s contributed to the extensive loss of body hair .
Behavioral hypothesis
In 1981, the anatomist C. Owen Lovejoy derived the bipedia from the social behavior of the early male great apes. His hypothesis stated that the bipedia arose as a result of a monogamous way of life: the males of the great ape species that developed into the early hominini had become monogamous and left their families alone during the day to look for food. They should have carried this food to their families, and the most effective way of getting around in this situation was to walk on two legs. Lovejoy's hypothesis, which, due to the lack of fossil records, cannot be directly substantiated by paleoanthropological findings, immediately provoked massive criticism. In particular, it was pointed out that monogamous primates usually do not have sexual dimorphism ; However, males of Australopithecus afarensis weighed almost twice as much as females - a characteristic one would expect in polygamous species. Furthermore, monogamous primates are strongly territorial, so they do not live in larger social groups; however, fossil evidence indicated that Australopithecus afarensis lived in groups.
In 2009, C. Owen Lovejoy revoked his hypothesis, referring to the finds of Ardipithecus ramidus in Hadar ( Ethiopia ): “Since the time of Darwin, the African great apes living today have mostly been the godfathers when early human evolution has been reconstructed. These models illustrate basic human behaviors as enhancements of behaviors seen in chimpanzees and / or gorillas (e.g., upright feeding posture, male dominance, tool use, culture, hunting, and warfare). Ardipithecus essentially falsifies such models, for the African great apes living today are highly derived relatives of our last common ancestor. "

Wat hypothesis
According to the so-called wading hypothesis (also: “bank hypothesis”), the hominini's bipedia developed as a result of the two-legged wading in search of food - for example for mussels - in waters with low water levels; two-legged wading is occasionally seen in bonobos , lowland gorillas, and proboscis monkeys . Two-legged wading has the advantage of keeping your head above water to breathe. In several publications, evolutionary biologist Carsten Niemitz tried to prove that none of the other explanatory models could plausibly explain the emergence of the upright gait. Parts of this theory can also be found in the water monkey theory .
Spine and pelvic girdle

The development of the ability to permanently walk upright required a large number of adaptations of the skeleton, particularly in the area of the spine , the pelvic girdle and hip joints , the knee joints , the feet and the foramen magnum (the large occipital opening at the base of the skull , where the spinal cord and brain merge). Changes affected both the structure of the bones and the skeletal muscles as well as their origin and insertion .
A permanent two-legged method of locomotion and standing upright both require that the center of gravity of the body is exactly vertically above the stepping surface of the feet. Therefore, the foramen magnum shifted forward during the formation of the upright gait and is now (like the pelvis, knees and feet) in the area of the body's center of gravity. If four-footed animals stand up on their hind legs, they only meet this requirement if their hind legs are bent at the hip and knee joints, as otherwise they would tip over forwards if the legs are stretched and vertically aligned. In the course of the evolution of the ancestors of anatomically modern humans, the body's center of gravity was therefore shifted towards the back. The development of the forward curvature of the spine ( lordosis ) in the area of the cervical spine and the lumbar spine contributed in particular to this , which is why the latter is particularly pronounced in pregnant women due to the shift in focus during pregnancy. At the level of the chest and in the area of the sacrum , on the other hand, the spine is curved backwards ( kyphosis ) and gives space to the internal organs, so that the typical impression of a double S-shaped curvature results (in predominantly four-footed monkeys it is C-shaped ). At the same time, the pelvic bones took on the additional task of supporting the internal organs.
Another special feature is the very narrow pelvic canal in comparison with all other human species , through which the fetus must pass at birth. Its internal dimensions are almost identical to the head size of the newborn, which means that “mother and newborn carry a considerable risk of a traumatic birth”.
The hypothesis, repeatedly advocated by researchers, that the pelvic girdle and pelvic canal remained relatively narrow because a wider pelvic belt would have resulted in greater leg spacing and this was disadvantageous for rapid movement, is controversial.


hands
Compared to other apes, the anatomically modern human's hands have a shorter palm, shorter fingers, and longer, stronger and more agile thumbs . In particular, the opposable thumb simplifies access and facilitates, for example, the skillful handling of tools. Responsible for the fact that the thumb - unlike other monkeys - can touch the tips of the other fingers with its tip, is a specially shaped thumb base joint . This also allows people to clench their hands into fists , which in turn makes the hand an effective punching tool (while boxing ) without exposing the fingers to excessive risk of injury. Even the orientation of the fine bone trabeculae (trabeculae) in the metacarpal bones of Australopithecus africanus has - on features that were interpreted as an indication of opposability of the thumb - two to three million years ago.
With the help of 39 test persons, the influence of the manufacture of stone utensils - the movements required for this - and the use of stone utensils on the evolution of the hand was also verified experimentally. Accordingly, the forces involved in cracking nuts were not great enough to significantly change the anatomy of the hand, but the preparation of tees when creating stone artifacts and the forceful slamming when opening long bones to get to their bone marrow.
Skull and brain development
In addition to the whales - especially the dolphins - the primates are considered to be the mammals with the most complexly developed brains . The growth of the brain is already greater in the human fetus (i.e. already in the womb) than in the fetus of the chimpanzee. A comparative genome analysis showed that the expression of genes and protein biosynthesis “in humans and chimpanzees differ dramatically, especially in the brain”, while the expression patterns in the liver and blood hardly differ from one another.
Around 300,000 years ago, the brain of early archaic Homo sapiens had the same volume as that of modern humans; Their rounded skull shape, on the other hand, developed much later: only fossils that are younger than 35,000 years old have the same round shape as today's humans.
Increase in volume

In the course of cerebralization, there was a tendency among hominini to increase the volume and surface area of the brain, especially in the area of the cerebral cortex , which among other things had considerable effects on food intake : the brain of anatomically modern humans makes up only about two percent of body weight off, but it consumes around 20 percent of the metabolic energy ; In the resting state, the basal metabolic rate of an adult corresponds to an output of around 100 watts , of which the brain accounts for 30 to 60 percent. With the increase in brain volume, there was a significant increase in the blood supply to the brain.
The reasons for the increase in the volume of the skull and for the diverse interactions between brain development, anatomical changes in the facial skull and other body features and, for example, the acquisition of food, are still relatively little researched. The enlargement of the brain of the hominini in the Australopithecine epoch around 3 to 2 million years ago and in Homo erectus , however, correlated with a cooling of the climate in Africa; Two calculations presented in 2000 and 2009 provided evidence that in cooler climates the heat radiation of the brain in the closed skull capsule proceeded more easily than before and therefore a "growth brake" was not applied, so that a larger and therefore more heat-producing brain developed could. A reduction in the mean temperature by 1.5 degrees Celsius may be sufficient for this. An alternative explanation assumes that the brain enlargement correlates with the population size and the necessary social assertiveness against competing conspecifics in a growing population was a driving force of brain development.
It is also noteworthy that the brain of people living today is roughly three to four percent smaller than the brain of ancestors 10,000 to 15,000 years ago. It is conceivable that this is a consequence of the warming after the end of the last large-area icing; however, it has also been argued that connectivity may have been optimized or that an optimized social structure improved the individual's chances of survival and this had an effect on brain size.
| Taxon | Brain volume in cm 3 |
|---|---|
| Chimpanzees | about 400 |
| Gorilla man | 535 |
| Sahelanthropus tchadensis | 365 |
| Ardipithecus ramidus | 280-350 |
| Australopithecus afarensis | 458 (335-580) |
| Australopithecus africanus | 464 (426-502) |
| Homo rudolfensis | approx. 750 (501–950) |
| Homo habilis | 610 (544-674) |
| Homo ergaster | 764 (640-888) |
| Homo erectus | 1003 (956-1051) |
| Homo antecessor | about 1000 |
| Homo heidelbergensis | 1204 (1130-1278) |
| Neanderthals | 1426 (1351-1501) |
| Homo floresiensis | 380 |
| homo sapiens | 1478 (1444-1512) |
The volume of the brain of the fossil hominini could be reconstructed at least approximately using skull fragments. The table opposite gives an overview of this. Since the brain has roughly the same density as water, the volume roughly corresponds to the mass in grams . The information on fossil species are estimates based on mostly incomplete and deformed finds; different textbooks can therefore contain different information.
The brain volume of the anatomically modern human has a considerable range , which is a direct consequence of the equally considerable variability of his body size. Since women are on average smaller than men, women also have a slightly smaller brain on average than men (see: Brains of men and women ).
In comparison with the various species of Australopithecines - in particular with Paranthropus aethiopicus , for example - all previously known species of the genus Homo have an extremely weak chewing apparatus with receding chewing muscles . According to US researchers working with Hansell H. Stedman, there is a temporal and functional relationship between this regression and the beginning of the brain's enlargement. Before about 2.4 million years, have mutation of a gene occurs that the protein MYH16 (myosin heavy chain 16) encoding ; In mammals, the MYH16 gene is only active in the jaw area - in the temporalis and masseter muscles - and causes the production of particularly resilient myosin chains in the muscle cells (see Heavy Chain ). The mutation has meant that this gene has since been inactive in the lineage that leads to people living today. It is no coincidence that the earliest fossils of the genus Homo come from the same geological epoch : The failure of the gene resulted in a marked reduction in the size of the individual muscle fibers and - as a result - the entire masticatory muscles. The associated loss of the considerable tensile force of muscles that attach to the skull and move the lower jaw was in turn a prerequisite for the brain, and with it the entire skull, to be able to enlarge later. Chimpanzees and other primates still have the intact, non-mutated gene.
Special adjustments
So far, there are only a few reliable findings about which regions of the brain were subject to a certain selection pressure in which epoch and therefore changed. In the resulting Endocast the so-called child of Taung (a three to four year old Australopithecus africanus ), however, remained a "suture" (ridging suture) received the forehead skull between the two halves, which - as in most cases at the same age children of Homo sapiens - even was not ossified ; In chimpanzee boys of the age of the Taung child, however, this seam is already ossified. From this it was concluded, among other things, that even in Australopithecus africanus the postnatal increase in brain volume - similar to Homo - was more pronounced than in the chimpanzees.
Three-dimensional reconstructions showed, for example, that the olfactory bulbs of anatomically modern humans take up about 12 percent more volume than those of Neanderthals and that in Homo sapiens the temporal lobe is also larger than it was in Neanderthals. The same applies to the upper parietal lobe , the increase in volume of which is also the cause of the more rounded parietal bone in Homo sapiens .
According to a study from 2016, the blood flow to the brain - except for Homo heidelbergensis and the Neanderthals - increased disproportionately in relation to the enlargement of the brain of the respective species.
While the brain volume of chimpanzees remains unchanged with age, it shrinks noticeably in aging people. This is presumably a consequence of the significantly increased life expectancy in humans compared to chimpanzees (approx. 80 to 90 years for humans, approx. 50 years for chimpanzees) and thus an evolutionary, relatively young phenomenon.
The loss of prominent bulges above the eyes and various other evolutionary changes in the total of 14 bones of the face - and the resulting changes in the muscle attachments - were brought into connection in a review article in 2019 with a more comprehensive non-verbal communication through minimal muscle movements, the indications of give subtle emotions and thus contribute to social interaction in groups.
Teeth and dentition

Teeth are highly resistant to damaging environmental influences, which is why they are the most common primate fossils found. Their size and shape, the thickness of their enamel and the ratio of the two stable carbon isotopes 12 C and 13 C in the enamel (see isotope analysis ) can provide information about the phylogenetic affiliation of their former owners, their social behavior and their diet. However, it is precisely these tissues and the similarly resistant pine that are “particularly susceptible to homoplasia ” (for parallel, independent development) as a result of similar diets , which can limit their use for taxonomic issues.
The evolutionary trend towards the reduction of the size of the teeth, which was already evident in Homo erectus , was interpreted as a result of the use of stone tools , with the help of which the food was broken up, which in turn reduced the chewing load of the teeth, upper jaw and lower jaw.
Reduction of the canines
Humans and the other great apes belong to the Old World monkeys . Its earliest known representatives - such as Aegyptopithecus - wore two large, elongated canines in the upper jaw , which were sharpened by constant rubbing on each of the lower front teeth (by honing ). At the same time, the upper jaw had two tooth gaps ( diastema ) between the canines and incisors , in each of which one of the canine teeth of the lower jaw fit. In the course of hominization, the canines became smaller and the tooth gap in the upper jaw disappeared.
As early as 1871, Charles Darwin recognized the chimpanzees and gorillas still living in Africa as the closest relatives of humans and therefore suspected that anatomically modern humans had also developed in Africa. At the same time, Darwin had interpreted the physique of these great apes in terms of an original feature . According to his guess
- “Man's earlier male ancestors were probably provided with large canine teeth; in proportion, however, as they gradually acquired the skill, stones, clubs or other weapons in the battle with their enemies to use, they are also their jaws have become less and less needed and teeth. In this case the jaws in connection with the teeth will have been reduced in size, as we can probably assume with certainty after countless analogous cases. "
Darwin's conjecture influenced countless attempts for more than 100 years to reconstruct the original dentition of the hominini - derived from the dentition of the chimpanzees. However, hypotheses about the teeth of the early hominini that can be substantiated by fossil finds were possible for the first time after the 4.4 million year old fossil Ardi and numerous teeth from other individuals of the species Ardipithecus ramidus had been discovered. Ardi neither had a particularly pronounced snout , nor did her male conspecifics have the dagger-like elongated canine teeth (" fangs ") known from chimpanzees and clearly visible from the outside . If Ardipithecus ramidus was rightly placed in the line of evolution of the hominini, its anatomical features would prove that the evolutionary trend towards smaller canines had already started more than four million years ago, long before the manufacture of the first stone tools , and thus the whole Pliocene lasted away.
The dagger-like canine teeth of the upper jaw of the male monkeys living today also regularly serve as a weapon in battles for hierarchy within their own group and in battles with individuals from other groups. The early onset of the reduction in size of the canines therefore suggests that due to sexual selection , the agonistic behavior and the imposing behavior also changed "long before the hominini had an enlarged brain and used stone tools."
Changes in the number of teeth and the snout

A second, long-term evolutionary trend associated with the reduction in the size of the teeth and the snout can be reconstructed from the reduction in the number of teeth. It is now widely recognized that the original higher mammals had three incisors , one canine , four front teeth, and three rear molars in each half of the jaw . Their tooth formula is therefore 3 • 1 • 4 • 3, their number of teeth was 44. All old world monkeys living today, however, have the tooth formula 2 • 1 • 2 • 3 and thus 32 teeth. In humans, the trend towards reduction can be observed directly insofar as the third (rear), so-called wisdom tooth, shows considerable differences in shape and time of eruption, and occasionally its tooth anus is completely absent; it can therefore be called a rudiment .
The Hominisation has also decreased in the course the number of roots with which the front molars are anchored in the jaw: The early types of Hominini from the group of australopithecines had two roots per front molar, Homo sapiens has only one root. In comparison to the chimpanzees, the change from deciduous to permanent dentition is delayed in humans during individual development , which is also an indication of a prolongation of the youth phase in humans. In the non-human great apes the eruption of permanent teeth begins at the age of 3.0 to 3.5 years, in humans, however, usually only at the age of 6; The cause is delayed formation of the tooth roots. Another characteristic of the young Homo sapiens is that the molar M1 and the incisor I1 of the permanent dentition break through together, followed by the incisor I2, while in the young chimpanzee only the molar M1 breaks through, followed by both incisors.
The shortening of the lower jaw, which Charles Darwin traced back to changes in eating habits, was confirmed in 2011 by a study of eleven populations living today. According to her, hunters and gatherers have longer and narrower jaws than sedentary people who feed on crops , dairy products or farmed animals .
Hairiness

What is striking about people living today is that they - as the only one of all primates living today - have extremely little body hair in addition to the mostly inconspicuous vellus hair . Since no paleoanthropologically relevant hair finds have been discovered so far, no statements can be made about the hair of extinct species of the hominini, nor can the period of time during which the fur was lost in its ancestors can be paleoanthropologically proven. The growth of the main hair, which has continued unchecked for a few years, also finds no parallel among primates; a clear function is not evident.
Hypotheses at the time
From molecular biological analyzes of the evolution of the human lice family (Pediculidae) and the time when dark skin color developed , some clues as to when the reduction in body hair occurred approximately.
The human lice family consists of the genera Pediculus and Phtirus . The pediculus includes the clothing louse adapted to humans ( Pediculus humanus humanus , in contrast to the head louse Pediculus humanus capitis also known as the "body louse " Pediculus humanus corporis ) and the species Pediculus schaeffi, which is adapted to chimpanzees . Phtirus occurs in humans as pubic louse ( Phtirus pubis ), in gorilla as Phtirus gorillae . From genetic analyzes it was deduced that the last common ancestor of Pediculus humanus spec. and Pediculus schaeffi existed about 6 million years ago and the last common ancestor of Phtirus pubis and Phtirus gorillae about 3 to 4 million years ago; also that the separation of Pediculus humanus humanus and Pediculus humanus capitis occurred 72,000 ± 42,000 years ago. From this the following time sequence can approximately be reconstructed: The ancestors of chimpanzees and humans and with them the ancestors of the parasites Pediculus schaeffi and Pediculus humanus spec have lived for 5 to 6 million years . separated from each other. Around 3 million years ago, the common phtirus ancestor, originating from the gorilla, was able to settle permanently with the ancestors of today's humans living at that time, with the simultaneous presence of Pediculus humanus spec . According to this, different hair biotopes probably already existed in the ancestors of humans in this epoch, for example on the head and in the pubic area, which were separated by zones with little hair (which would roughly correspond to our current hair growth). Finally, about 60,000 to 80,000 years ago, the splitting of Pediculus humanus spec. into the two subspecies Pediculus humanus capitis and Pediculus humanus humanus as signs of the use of clothing. A second, independent study confirmed this argument in principle, but came to higher values: According to it, the latter split occurred at the latest 83,000 years ago, but possibly also 170,000 years ago.
Information about the approximate point in time was also provided by calculations of the time when dark skin color appeared. Many researchers suspect that the hairy early species of the hominini - like the chimpanzees - had pink skin. A specific variant of the MC1R gene, which is involved in skin pigmentation, is found in all dark-skinned Africans and has existed for 1.2 million years , according to a study by researchers at the University of Utah . From this it was in turn deduced that the body hair must have been quite thin back then.
Hypotheses on causes and consequences
Evolutionary biologists differ on the question of why the fur was reduced. “Perhaps the most plausible theory assumes that nakedness, together with the multiplication of sweat glands, originally served to regulate body temperature. The feature would have arisen in early humans ( H. erectus ) around two million years ago as an adaptation to persistent running under heat stress. ”This is supported by the fact that humans have much more eccrine sweat glands than their hairy relatives and therefore - due to lack of fur - the The thin sweat produced by them can evaporate directly on the skin ( "evaporation cold" ). This hypothesis is supported, among other things, by a model calculation, according to which only a largely hairless individual of the genus Homo , capable of profuse sweating, could have been active during the day in hot, open terrain. This was an evolutionary advantage that made endurance hunting possible while potentially reducing susceptibility to ectoparasites . A mutation in the EDAR gene probably caused the inhabitants of East Asia around 30,000 years ago to have the density of their sweat glands even greater and their hair thicker at the same time.
One consequence of the reduced hairiness was that the now largely unprotected skin was exposed to intense African sunlight: “In order to keep out the harmful UV radiation , the body 'invented' the production of melanin and thus the darker skin color, which made it possible for people to to be able to survive at all in these latitudes . ”Among other things, pigmented skin prevents the body's folic acid supply from being impaired by excessive UV radiation. According to calculations by Nina Jablonski (California Academy of Sciences), skin color can be changed relatively easily, so that a change from light to dark or from dark to light can take place within just 100 to 200 generations. Therefore, the skin color is "worthless for determining the phylogenetic relationship" of populations living today.
Another consequence of the reduced hairiness was that the skin that had become visible had to be protected more strongly than before by cornification : “Comparisons of the human and the chimpanzee genome showed that we differ from our closest relatives in those genes whose proteins have skin properties In addition, the coloring of the skin could become a social signal and in anatomically modern humans it could also be used for intra-species communication, as emotional states such as fear or anger are accompanied by pale or blushing . In addition, signals about emotional states shifted from bristling hair to complex facial expressions .
It is also "obvious appearance and slow growth of pubic hair in puberty , the sexual maturity signaled."
Social behavior
The paleoanthropologist Richard Leakey described the social behavior of man's ancestors who lived seven million years ago as follows: “Instead of living as family groups in nomadic associations like today's hunters and gatherers , the first humans probably lived like robed baboons . At that time, packs of around 30 individuals roamed in a coordinated manner through an extensive area and returned at night to their preferred sleeping places in steep cliffs or groups of trees. Adult females and their offspring made up the majority of the pack, while sexually mature males were only present in small numbers. "
Our current social behavior , which manifests itself for example in empathy , social closeness / distance , willingness to aggressive behavior , fearfulness or in the reactions to stressors , is influenced to a considerable extent by hormones such as adrenaline , oxytocin , serotonin and testosterone . According to a study published in 2018, humans differ from all other primates. a. due to a "dramatic" increase in dopamine release in the striatum - also in comparison with the other great apes - and a lower release of acetylcholine . The production of serotonin and neuropeptide Y in the striatum is also increased. This combination of increased and decreased hormone production "favors externally controlled behavior and considerably increases the feeling for social signals that promote social adaptation, empathy and altruism ."
childhood
The very narrow pelvic canal in female Homo sapiens - in relation to the body size of all other human species - is particularly noteworthy insofar as the brain size of a newborn in Homo sapiens is only around 28 percent of the brain size of an adult. A chimpanzee, on the other hand, is born with 40 percent the brain size of its adult individual, and the early Homo erectus was also born in the Old Pleistocene - more than a million years ago - probably around 35 percent the brain size of an adult, although its pelvic canal is already significantly larger was than that of Australopithecus afarensis . Macaques are even born with 70 percent of the brain volume of adult conspecifics.
In comparison with Homo erectus , the growth in size of the brain in Homo sapiens is also slower in the months after birth. This was derived from a child's skull discovered in Java in 1939 ("Mojokerto child"; archive number Perning I), which was dated to 1.8 million years ago in 1994 using the 40 Ar- 39 Ar method . In 2004, the child was assigned an age at death of approximately one year and a brain volume of approximately 70 to 90 percent of an adult; For comparison: children of Homo sapiens at the age of one have only about 50 percent of the brain volume of an adult.
Two evolutionary trends can be derived from this for the line of development that leads to people living today: On the one hand, the pelvic canal enlarged, which suggests the passage of a brain that has grown larger and larger even before birth; on the other hand, the brain - in relation to its size in adults - became smaller and smaller at birth. The brain growth was thus postponed to a considerable extent in the time after birth, which resulted in a significant extension of childhood and the extended social care associated with it by adults 160,000 years ago: the phase of childhood, youth and adolescence - the is at the same time a phase of upbringing and intensive social learning - lasts twice as long in anatomically modern humans as in chimpanzees and led to a lengthening of the interval between two births. In comparison with Homo erectus and with the Neanderthals, the speed of development of a child of Homo sapiens was much slower.
Life expectancy
| Taxon | Duration of gestation (days) |
Life expectancy (years) |
|---|---|---|
| Lemurs | 120-135 | 14-15 |
| Macaques | 165 | 27-28 |
| Gibbons | 210 | 30-40 |
| Chimpanzees | 228 | 40-50 |
| People | 266 | > 70 |
One of the special features of primate evolution in terms of reproductive biology is that not only childhood, but also the prenatal phase (the length of gestation ) and overall life expectancy have increased over the course of the tribal history (see table).
The tendency towards a longer life span was probably associated with special genetic adaptations to ward off potential pathogens. This was, among others, derived from a study on the functioning of the apolipoprotein E . Its variant ApoE3, which is only present in humans, makes a significant contribution to the fact that humans are less sensitive to inflammation and infections than, for example, chimpanzees.
Another consequence of the increased life expectancy - it is often argued - was the emergence of menopause : “In order to increase the likelihood of surviving the death of the mother for the offspring of mothers, new reproductive strategies developed, ie older women do not give birth still children themselves, but invest in caring for the offspring of their own daughter. The ' invention of the grandmother ' presumably represents the functional ultimate cause of female menopause. ”In fact, it has been proven for Finland and Canada that a particularly long life after menopause correlates with a particularly large number of grandchildren .
sexuality
In all mammals, sexual behavior consists of a sequence of actions and reactions by the sexual partners , each of which "very specifically reinforces one another. […] This means that among other things the sexual behavior of these animals is not 'instinctive', that is, not exclusively determined by themselves. ”Rather, the sexual behavior of these animals - and especially the great apes -“ is to a high degree of practice and Experience ”. “Humans are innate with certain basic sexual responses, but they are not specifically 'programmed' to mate. They are therefore almost entirely dependent on observation and experience. Your sexual behavior is extremely variable ”.
While emotions such as fear and fear as well as sadness and melancholy can already be influenced by pharmaceutical therapies, the neurobiological and physiological basis of the emotion love is still largely not understood.
Genetically determined peculiarities
- Constant willingness to conceive outside of pregnancy
- no observational traits for the sexual cycle
- Primary reproductive organs hidden from direct view in the adult female
- Sexual intercourse largely without synchronization with ovulation
- Formation of breasts in adult women regardless of lactation , the size of which is determined by adipose tissue, not by glandular tissue; the striking coloration of the nipple region and the size of the areolas are also unique among mammals.
- The onset of fertility is considerably delayed compared to other (also long-lived) primates
- End of fertility in women long before they die: From the age of around 50, women experience the end of their fertility due to menopause
- Highlighting of the adult primary genital organs by clearly separated pubic hair with otherwise little hair
- Loss of penile bone
- Formation of an extremely large penis for primates in the erect state , which cannot be hidden (without aids)
Culturally established features
- Monogamy : In many ethnic groups there are more or less long-term relationships between a woman and a man, which by no means excludes a tendency for both sexes to 'flirt' . But there are also populations in which polygyny or polyandry occur and are officially tolerated. Genetic research suggests that monogamy was the common form of couple relationship for long periods of time in the ancestors of modern humans. This also meant that the men were jointly responsible for the survival of their children in the long term and directly.
- Regulations and prohibitions of sexual practices in many (or most) cultural and religious regulations.
- Coupling of Shame and Sexuality: Humans are the only species that can develop shame for sexuality; Sexual intercourse usually takes place in camera, while animals usually copulate in front of their conspecifics. A number of authors take the position that hidden ovulation, sexuality for pleasure, and the privacy of the sexual act are features that have favored the bond between a man and a woman, that is, the long-term economic cooperation of couples necessary to raise children .
Speaking ability

The ability to articulate extremely complex distinguishes anatomically modern humans from all other great apes . When the speaking ability based on this developed and what selection pressure caused this is largely unknown due to a lack of fossil evidence. It is possible that the complex muscle movements required for speaking (fast and coordinated movements of the lips , lower jaw and tongue ) initially served facial expressions , i.e. visual communication .
The prerequisite for the development of the ability to speak was, among other things, the development of a vocal tract unique among primates and its motor control as well as the mental ability to generate an infinite number of names with a finite number of sounds by using the sounds in a certain sequence ( syntax ). "Compared to the great apes , the larynx, which is important for speech production, is lower and is also constructed differently in important details, such as the vocal cords ." In particular, the free mobility of the tongue contributes to the fact that a particularly wide variety of sounds is generated can be.
Since the discovery of the hyoid bone of a Neanderthal man in the Israeli Carmel Mountains in the mid-1980s, it has been considered certain that the anatomical prerequisites for linguistic communication were also given in the Neanderthal man. However, it is unclear whether this also applies to the last common ancestor of Neanderthals and Homo sapiens . The significance of a mutation in the gene coding for the forkhead box protein P2 - which has been assigned the role of a “language gene” in the mass media - is also unclear. According to a paleogenetic study, it has existed for 200,000 years in the form that is detectable in humans today.
In tribal history, however, the ability of great apes to form combinations of symbols and communicate them to other individuals is much older. This is what Roger Fouts' research on chimpanzees has shown since the 1970s :
“So they have the auditory and cognitive abilities to 'understand' language even though they cannot speak themselves. From these experiments it can be concluded that syntactic and symbolic understanding has evolved in other areas of behavior, namely in social cognition. There is no fundamental difference between humans and non-human primates. "
The main difference between human language and the utterances of other animals is the grammar , which makes it possible to represent complex relationships. People are only able to spontaneously fully learn the grammar of a language and to learn to speak it without an accent in the sensitive phase of toddlerhood. Small children who grow up in an environment in which no fully developed language is used for communication - for example in situations in which a pidgin is used for communication - spontaneously transform it into a language with all grammatical functions - a so-called creole language . Steven Pinker derived the idea of a "language instinct" from this, which should, to a large extent, dictate the characteristics of natural languages. In contrast, great apes are able to learn individual words and put them together to express new meanings, but they cannot build grammatically correct sentences to represent more complex contexts of meaning.
One hypothesis supported by many researchers is that communication through sounds has gradually replaced social grooming and, like this, contributed to the cohesion of the group, that is, the ability to speak emerged “in a highly social, potentially cooperative context, linked and with at least three characteristics: mutual attention , common intentions and theory of mind . "
When, among other things, based on these abilities, the anatomical changes in the vocal tract and the formation of today's language center , verbal communication developed into symbolic language, can only be clarified according to the current state of knowledge, “if we have unmistakable evidence from the archaeological evidence for have syntactic and symbolic linguistic skills at a high level. These qualifications were probably not yet achieved at the homo erectus stage. "
Culturally traditional characteristics
In humans, culturally traditional characteristics include historicity and morality as well as the passing on of knowledge through language (i.e. through symbolic communication ), the much more complex cooperation with the increasing group size , the production of works of art and the use of technology . Processes of cultural evolution have been researched under the heading of cultural ethology since the 1970s .
In 1976 , the evolutionary biologist Richard Dawkins proposed the concept of memetics for information that is disseminated and multiplied ("replicated") solely by human consciousness . A counterpart to the gene was thus designed, the basic idea being that certain information (“memes”) replicate because of their advantageous nature and prevail over other information, just like the “successful” genes do. In this sense, traditions of tool manufacture ( example: hand ax ), work processes ( example: fire , agriculture ) or rituals of early hominids can be referred to as memes or memplexes ("meme complexes"), which replicate from brain to brain due to their beneficial properties . Such a model can be useful, for example, to understand the dominant right-handedness of today's people as a result of a memetic transmission of toolmaking and other work processes.
However, anatomically modern humans are not the only species that pass on learned properties to the next generation. A well-known example is the “potato washing” of a population of Japanese macaques on the island of Kōjima . In a review by Andrew Whiten and Jane Goodall in 1999, several dozen group-specific traditional behaviors were found in chimpanzees, including the use of tools in chimpanzees . In 2001, for example, "Culture in whales and dolphins " was described, in 2003 in orangutans and white-shouldered capuchins . An essential difference between animals and humans is therefore not in the existence of culturally traditional characteristics, but in the fact that young animals acquire the behavior of their parents through observational learning, while humans also gain their knowledge more actively through instructions ('teacher' and 'pupil'). pass on.
Spoken language
The greatest genetic diversity of humanity was in African populations of the south Sahara proven. In a similar way as the genetic and - connected with this - the phenotypic diversity decreases with distance from Africa, since additional populations in the non-African world were mostly founded by only a few individuals, the number of phonemes used also decreases . From this it was deduced that the origin of the first traditional human languages (the origin of the language ) was in West Africa , Central Africa or in southern Africa.
There are numerous hypotheses about the structure of the first languages - so-called proto languages . For example, it is assumed by some researchers that individual words were initially used without any arrangement in syntactic structures (without arrangement in “ sentences ”); There is consensus in paleolinguistics that spoken words initially referred primarily to objects and events , and not to ideas . The noticeable accumulation of tonal languages , in which a change in the pitch or the tone course in a syllable is usually accompanied by a change in the meaning of the corresponding word (or the morpheme ), in tropical and subtropical regions is explained by the hypothesis that such Speeches could not have developed in a less humid environment, since dry air made it difficult for the larynx to hit the right note.
After analyzing 2,200 living languages and dead languages published Murray Gell-Mann and Merritt Ruhlen 2011 a hypothesis to the original sentence structure. Accordingly, the majority of living languages arrange their parts of the sentence in the order subject - verb - object , that is: "I - see - lion"; Most dead languages, however, arranged the parts of the sentence in the order subject - object - verb , "I - lion - see". According to Gell-Mann and Ruhlen, all 'subject-verb-object languages' can be derived from 'subject-object-verb languages', but there is not a single indication of an opposite change in the sentence structure. This was interpreted by Merritt Ruhlen as proof that "the presumed original language had a 'subject-object-verb word order'."
The oldest drawn symbols - for example circles, spirals, points, lines, open and closed triangles and hands - are around 35,000 years old, distributed over the African and European settlement area of Homo sapiens ; however, it is unclear whether these symbols can be interpreted as early evidence of the beginnings of a written language .
Tool use

For a long time, the use of tools was seen as an important definition criterion to distinguish the genus Homo from the Australopithecines. This only changed after Jane Goodall demonstrated tool use in animals in 1964 . It is assumed that - comparable to the chimpanzees living today - also processed pieces of wood were used by australopithecines, which, however, cannot be archaeologically proven due to their transience.
In Kenya, stone tools ( rubble tools ) that are 3.3 million years old were discovered at the Lomekwi site , which in 2015 were called the Lomekwian type by their discoverers . The oldest stone tools of the Oldowan type are up to 2.6 million years old and thus also older than the earliest representatives of the Homo genus . They were found in connection with bones on which cutting traces from the processing with stone tools have been preserved and which are therefore considered to be relatively safe. From today's sources, the author of these tools is Australopithecus garhi . 3.4 million year old carvings on two wild animal bones from Dikika were also traced back to stone tools and attributed to Australopithecus afarensis ; From the same find layer , however, no stone utensils are known so far, which is why the interpretation of the incisions as cutting marks is controversial.
In South Africa discovered 500,000 year old stone artifacts were in 2012 as projectile points interpreted; If this interpretation is correct, the finds from the site Kathu Pan 1 (KP1) would be the oldest evidence of the use of spears.
A convincing correlation between biological development and the presumed selection advantage through the use of simple rubble devices has not yet been proven. Instead, is ecological niches as possibly the most important driving force behind due to the very different regional food supply speciation viewed. This process began at least 3 million years ago, with the omnivores (generalists) prevailing in the long term . For younger Homo sapiens populations, on the other hand, it is still considered likely that the imparting of special manufacturing techniques for stone utensils to the subsequent generation had an impact on social behavior and the ability to learn, among other things, and also promoted the ability to act according to plan.
At least 72,000 years ago, stones made of a certain silicified rock ( silcrete ), from which tools with sharp edges were to be made, were routinely placed in the fire; after heating could be the reductions from the lighter core to separate.
The domestication of the wolf was also interpreted to mean that it was shaped into a '(hunting) tool' for humans.
dress
488 flax fibers - including 58 allegedly dyed fibers - are considered the oldest evidence of the manufacture of clothing. They come from the Dzudzuana Cave in Georgia and were recovered in 2007 and 2008 from a layer of soil whose age was dated to 36,000 to 31,000 years (yr BP ka). Fiber finds - probably from nettles - from Dolní Věstonice ( Czech Republic ), which are 32,000 to 29,000 years old and also come from Cro-Magnon humans ( Homo sapiens ), are only slightly younger .
When the first pieces of clothing (probably made of fur) were made is unknown, as there are no older archaeological finds. However, genetic analysis of human lice has led to the conclusion that Homo sapiens wore clothes at least 70,000 years ago, possibly even 170,000 years ago. There is also speculation among experts that the much earlier spread of Homo heidelbergensis and the Neanderthals in northern Eurasia required “cold-resistant clothing”, “in order to survive even when temperatures fell below 10 degrees Celsius for months without permanent children and for small children persistent hypothermia infallibly fatal damage to remain able to act. "
nutrition
According to the current state of knowledge, anatomically modern humans are “naturally” neither a pure “ carnivore ” nor a pure “ herbivore ” (herbivore), but an “ omnivore ” (omnivore). For the earliest fossils assigned to the genus Homo , it has been proven that - similar to Australopithecus africanus and Paranthropus robustus - more than 50 percent of their daily caloric intake was nourished on C 3 plants and a further significant proportion on C 4 plants .
The use of tools later had a major influence on human nutrition and, as a result, also changed the shape of the bone plates of the head, the jaws and the teeth. If one converts the amount of time monkeys spend daily on food intake to human body mass, one would expect Homo sapiens to do 48 percent of daily activity; in fact, it is only just under five percent. In particular the smaller molars compared to Homo habilis and Homo rudolfensis - around 1.9 million years ago in Homo erectus and later in Neanderthals and Homo sapiens - can most likely be interpreted as a result of "processed" food. 1.95 million years old bone finds from Kenya testify, for example, that in addition to antelope meat, the meat of numerous animals living in the water - including turtles , crocodiles and fish - was consumed. Similar to today's baboons , Australopithecus afarensis still fed on a predominantly plant-based, but not very hard-fiber diet, while the individuals of the genus Homo were already consuming increasingly protein-rich foods two million years ago - which in turn favored the gradual enlargement of the brain.

According to the current state of research, it is certain that Homo erectus was the first to learn to control fire , which is an exclusive ability of the Homo species among all living beings ; however, chimpanzees also seem to prefer cooked foods. The oldest undisputed site of burnt human food remains is Gesher Benot Ya'aqov in northern Israel , which is around 790,000 years old. In Europe, on the other hand, the oldest evidence of fireplaces dates from around 400,000 before today ( Schöningen in Germany and Beeches Pit in England), while older sites of hominine fossils such as the cave of Arago in France or the caves in the Sierra de Atapuerca (Sima del Elefante, Gran Dolina) show no traces of fire use; Despite temperatures of sometimes below zero degrees, Homo erectus was able to colonize Europe without generating heat from fireplaces. As a result, Homo erectus did not practice heating food until relatively late in Europe, whereas Homo sapiens has probably been a distinguishable species since the beginning of its existence.
Even today humans are “omnivores” (omnivores); What is disputed, however, is the proportion of food consumption in premodern societies that accounted for meat and vegetable food: While the Evenks in Siberia and the Eskimos mainly lived on meat , the peoples in the Andes lived primarily on vegetable foods; in the majority of the hunter-gatherer peoples still alive today, however, well over half of the diet comes from animals.
For men living in Europe 30,000 years ago, the average height was estimated at 1.72 meters; later - apparently as a result of the expansion of agriculture - the food situation deteriorated, so that the body size steadily decreased until 4,000 years ago. Another decrease in size occurred in the Middle Ages. The height of European males has only returned to its original value for around 100 years and has recently exceeded it.
Works of art

As the earliest evidence of symbolic communication apply with geometric engravings decorated ocher pieces from the Blombos Cave in South Africa , which have been dated at about 77,000 years. Pierced snail shells ( Nassarius gibbosulus ) from the Grotto des Pigeons near Taforalt ( Oujda region , Morocco ), which were found with adhering ocher residues, are still 5000 years older . About 60,000 years old are 270 fragments of ostrich eggs , which were also discovered in South Africa - in the Diepkloof cave - and also show geometric patterns ( hatching , parallel and crossing lines). With an age of less than 40,000 years, the cave paintings known from Europe as well as objects such as the lion people and the Venus from the Cave Rock are much younger .
Social care
Social care , that is, the altruistic care of individuals who are only partially able to act by other members of their group, is largely limited to the feeding and protection of young animals in the great apes living today. However, the characteristics of some fossils of the genus Homo have been interpreted to show clear indications of social welfare for the benefit of elderly adults. In Georgia , for example, an approximately 1.8 million year old, toothless skull with a corresponding edentulous lower jaw was discovered among the hominine fossils of Dmanissi , whose "healed" tooth sockets allow the conclusion that the individual - although he could no longer chew coarse food - with strongly shredded food and must have been socially integrated despite his disability.
The discovery of a roughly 500,000 year old Homo heidelbergensis from the Sierra de Atapuerca in Spain is interpreted in a similar way : The male spine shows signs of pronounced kyphosis (a "hump"), of spondylolisthesis and of Baastrup's disease , so that this individual is probably just himself with a "walking stick" could move upright. Because of these painful diseases, it was neither able to take part in the hunt nor to carry heavy loads, so that it was probably particularly dependent on the social care of its group.
Spirituality and religiosity

Charles Darwin had already suspected that man's inclination towards spirituality and religiosity only developed in the course of recent tribal history:
- "We have no evidence that man from his origins had the ennobling belief in the existence of an almighty God."
Darwin interpreted this tendency as a result of certain cognitive skills , i.e. ultimately made possible by the interconnection of the nerve cells in the brain :
- “As soon as the meaningful faculties of imagination, amazement and curiosity, in connection with a faculty of thought, have been partially developed, man will have sought to understand what is going on around him and will also become obscure about his own existence have begun speculating. "
Today's cultural anthropologists attribute the existence of spirituality and religiosity to the fact that they promote cooperative behavior between like-minded strangers and thereby contribute to the creation and maintenance of stable social groups; However, it is unclear whether this is to be interpreted in the sense of an evolutionary adaptation or - comparable to musicality - as a 'concomitant phenomenon' of other cognitive abilities.
The epoch in which these cognitive abilities first appeared is controversial: some researchers have established a connection with the oldest surely dated burials 95,000 years ago in Qafzeh , Israel ; others with the oldest certainly dated works of art such as Venus vom Hohlefels around 35,000 years ago.
The neurophysiological basis of mental abilities has so far been largely unexplored, but there are first indications at least with regard to spirituality; For example, Vilayanur S. Ramachandran pointed out: "Every medical student learns that patients with epileptic seizures that develop [in the left temporal lobe ] often have intense spiritual experiences during the seizures ..." Ramachandran also emphasized the uniqueness of such experiences:
- “We humans have many qualities that are unique to our kind, but none of them are as puzzling as religion - our tendency to believe in a higher power that transcends the world of appearances . It is extremely unlikely that any other creature can ask about 'the meaning of it all'. "
See also
literature
- Winfried Henke , Hartmut Rothe : Incarnation. Fischer Taschenbuch Verlag, Frankfurt am Main 2003, ISBN 3-596-15554-1 .
- Thomas Junker : The evolution of man. 3rd, completely revised edition. CH Beck, Munich 2018, ISBN 978-3-406-72276-9 .
- Alice Roberts : The Beginnings of Mankind. From the upright gait to the early high cultures. Dorling Kindersley Verlag, Munich 2012, ISBN 978-3-8310-2223-6 .
Web links
- Wildness within us - human evolutionary heritage. ( Memento from March 17, 2008 in the Internet Archive ) Lecture by Inge Schröder, November 1998. The author is a qualified anthropologist at the Kiel Science Center.
Individual evidence
-
↑ Stephen C. Stearns et al .: Measuring selection in contemporary human populations. In: Nature Reviews Genetics . Volume 11, 2010, pp. 611-622, doi: 10.1038 / nrg2831
Emmanuel Milot et al .: Evidence for evolution in response to natural selection in a contemporary human population. In: PNAS . Volume 108, No. 41, 2011, p. 17040–17045, doi: 10.1073 / pnas.1104210108
Jaleal S. Sanjak et al .: Evidence of directional and stabilizing selection in contemporary humans. In: PNAS. Volume 115, No. 1, 2018, pp. 151–156, doi: 10.1073 / pnas.1707227114 -
↑ Katherine M. Kirk et al .: Natural selection and quantitative genetics of life-history traits in western women: a twin study. In: evolution. Volume 55, No. 2, 2001, pp. 423-435, doi: 10.1111 / j.0014-3820.2001.tb01304.x
Alexandre Courtiol et al .: Natural and sexual selection in a monogamous historical human population. In: PNAS. Volume 109, No. 21, 2012, pp. 8044-8049, doi: 10.1073 / pnas.1118174109 - ↑ Alexandre Courtiol et al .: The Demographic Transition Influences Variance in Fitness and Selection on Height and BMI in Rural Gambia. In: Current Biology. Volume 23, No. 10, 2013, pp. 884-889, doi: 10.1016 / j.cub.2013.04.006
-
↑ Sandra Wild et al .: Direct evidence for positive selection of skin, hair, and eye pigmentation in Europeans during the last 5,000 y. In: PNAS. Volume 111, No. 13, 2014, pp. 4832–4837, doi: 10.1073 / pnas.1316513111
Strong positive selection has changed the appearance of Europeans over the past 5,000 years. On: idw-online.de of March 10, 2014
Humans are still evolving - and we can watch it happen. On: sciencemag.org from May 17, 2016 - ↑ C. Owen Lovejoy : The Origin of Man. In: Science . Volume 211, 1981, pp. 341-350, doi: 10.1126 / science.211.4480.341 - "Five characters separate man from other hominoids - a large neocortex, bipedality, reduced anterior dentition with molar dominance, material culture, and unique sexual and reproductive behavior. "
- ↑ Chris Stringer : Evolution: What makes a modern human. In: Nature . Volume 485, 2012, pp. 33-35, doi: 10.1038 / 485033a
- ↑ Literally: "large, linear bodies, elongated legs, large brain sizes, reduced sexual dimorphism, increased carnivory, and unique life history traits (eg, extended ontogeny and longevity) as well as toolmaking and increased social cooperation." Quoted from: Susan C. Antón, Richard Potts and Leslie C. Aiello: Evolution of early Homo: An integrated biological perspective. In: Science. Volume 345, No. 6192, 2014, doi: 10.1126 / science.1236828
- ↑ Nina G. Jablonski : Human Skin Pigmentation as an Example of Adaptive Evolution. In: Proceedings of the American Philosophical Society. Volume 156, No. 1, 2012, pp. 45–57, full text (PDF) ( Memento from April 30, 2015 in the Internet Archive )
- ^ Rolf M. Quam et al .: Early hominin auditory ossicles from South Africa. In: PNAS. Volume 110, No. 22, 2013, pp. 8847-8851, doi: 10.1073 / pnas.1303375110
- ↑ Carol V. Warda et al .: Thoracic vertebral count and thoracolumbar transition in Australopithecus afarensis. In: PNAS. Volume 114, No. 23, 2017, pp. 6000–6004, doi: 10.1073 / pnas.1702229114
- ↑ a b Manuel Will and Jay T. Stock: Spatial and temporal variation of body size among early Homo. In: Journal of Human Evolution. Volume 82, 2015, pp. 15-33, doi: 10.1016 / j.jhevol.2015.02.009
- ↑ a b For an overview see: WEH Harcourt-Smith, Leslie C. Aiello : Fossils, feet and the evolution of human bipedal locomotion. In: Journal of Anatomy . Volume 204, No. 5, 2004, pp. 403-416, doi: 10.1111 / j.0021-8782.2004.00296.x , PMC 1571304 (free full text).
- ↑ C. Owen Lovejoy: Evolution of Human Walking. In: Scientific American . November 1988, pp. 118-125
- ^ Richard Leakey : The origin of human kind. Phoenix, a division of Orion Books Ltd., 1995, p. 13.
- ↑ www.pbs.org: The Transforming Leap, from Four Legs to Two. An overview of the state of research.
- ↑ Winfried Henke , Hartmut Rothe : Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, p. 55
- ↑ Peter J. Fernández, Carrie S. Mongle, Louise Leakey et al .: Evolution and function of the hominin forefoot. In: PNAS. Volume 115, No. 35, 2028, pp. 8746-8751, doi: 10.1073 / pnas.1800818115 .
- ^ Mary Leakey et al .: Pliocene footprints in the Laetoli beds at Laetoli, northern Tanzania. In: Nature. Volume 278, 1979, pp. 317-323, doi: 10.1038 / 278317a0
-
↑ David A. Raichlen et al .: Laetoli footprints preserve earliest direct evidence of human-like bipedal biomechanics . In: PLoS ONE . Volume 5, No. 3: e9769. 2010; doi: 10.1371 / journal.pone.0009769 .
Prehuman with crossed cross. On: Wissenschaft.de from March 20, 2010. - ↑ Robin H. Crompton et al .: Human-like external function of the foot, and fully upright gait, confirmed in the 3.66 million year old Laetoli hominin footprints by topographic statistics, experimental footprint formation and computer simulation. In: Journal of the Royal Society Interface. Volume 9, No. 69, 2011, pp. 707-719, doi: 10.1098 / rsif.2011.0258 .
- ^ Vivek V. Venkataraman et al .: Tree climbing and human evolution. In: PNAS. Volume 110, No. 4, 2013, pp. 1237–1242, doi: 10.1073 / pnas.1208717110 , full text (PDF)
- ↑ Did Lucy walk, climb, or both? Australopithecine ancestors - arboreal versus terrestrial habitat and locomotion. On: eurekalert.org of December 31, 2012
-
^ Leoni Georgiou et al .: Evidence for habitual climbing in a Pleistocene hominin in South Africa. In: PNAS. Advance online publication of March 30, 2020, doi: 10.1073 / pnas.1914481117 .
Evidence of fossil hominin locomotion. On: eurekalert.org from March 30, 2020. - ^ Carol V. Ward, William H. Kimbel, and Donald C. Johanson : Complete Fourth Metatarsal and Arches in the Foot of Australopithecus afarensis. In: Science. Volume 331, No. 6018, 2011, pp. 750-753, doi: 10.1126 / science.1201463
- ^ "The Ileret prints show that by 1.5 Ma, hominins had evolved an essentially modern human foot function and style of bipedal locomotion." Matthew R. Bennett et al .: Early Hominin Foot Morphology Based on 1.5-Million-Year-Old Footprints from Ileret, Kenya. In: Science. Volume 323, 2009, pp. 1197-1201, doi: 10.1126 / science.1168132 . - In: New Scientist of May 23, 2009, p. 24, Richard Leakey pointed out that the assignment of the traces to Homo erectus is not mandatory, since Paranthropus boisei and Homo habilis also occurred at the same time and in the same African biotopes .
- ^ Dennis M. Bramble, Daniel E. Lieberman: Endurance running and the evolution of Homo. In: Nature. Volume 432, 2004, pp. 345-352, doi: 10.1038 / nature03052
- ^ Christopher L. Vaughan: Theories of bipedal walking: an odyssey. In: Journal of Biomechanics. Volume 36, No. 4, 2003, pp. 513-523, doi: 10.1016 / S0021-9290 (02) 00419-0 , full text (PDF; 990 kB)
- ↑ The cover of Science from June 1, 2007 contains a similar photo of a “standing” orangutan from Sumatra, see sciencemag.org
- ↑ SKS Thorpe et al .: Origin of Human Bipedalism As an Adaptation for Locomotion on Flexible Branches. In: Science. Volume 316, 2007, pp. 1328-1331, doi: 10.1126 / science.1140799 ; cf. on this: upright on the branch. On: sueddeutsche.de from May 17, 2010
-
↑ C. Owen Lovejoy et al .: The Pelvis and Femur of Ardipithecus ramidus: The Emergence of Upright Walking. In: Science. Volume 326, 2009, pp. 71, 71e1-71e6, doi: 10.1126 / science.1175831
C. Owen Lovejoy et al .: Careful Climbing in the Miocene: The Forelimbs of Ardipithecus ramidus and Humans Are Primitive. In: Science. Volume 326, 2009, pp. 70, 70e1-70e8, doi: 10.1126 / science.1175827
C. Owen Lovejoy et al .: Combining Prehension and Propulsion: The Foot of Ardipithecus ramidus. In: Science. Volume 326, 2009, pp. 72, 72e1-72e8, doi: 10.1126 / science.1175832 - ↑ quoted from an interview ("Sensationsfund Ardi - attraction instead of aggression") in: Süddeutsche Zeitung of October 2, 2009, p. 24, full text
-
↑ Tracy L. Kivella, Daniel Schmitt: Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor. In: PNAS. Volume 106, No. 34, 2009, pp. 14241-14246, doi: 10.1073 / pnas.0901280106
Bipedal humans came down from the trees, not up from the ground. On: eurekalert.org of August 10, 2009 - ↑ Madelaine Böhme et al .: A new Miocene ape and locomotion in the ancestor of great apes and humans. In: Nature , Volume 575, 2019, pp. 489-493, doi: 10.1038 / s41586-019-1731-0 .
-
↑ Kevin D. Hunt: The evolution of human bipedality: ecology and functional morphology. In: Journal of Human Evolution. Volume 26, No. 3, 1994, pp. 183-202, doi: 10.1006 / jhev.1994.1011
Kevin D. Hunt: The postural feeding hypothesis: an ecological model for the evolution of bipedalism. In: South African Journal of Science. Volume 92, 1996, pp. 77-90. - ↑ a b C. Owen Lovejoy: Reexamining Human Origins in Light of Ardipithecus ramidus. In: Science. Volume 326, 2009, pp. 74, 74e1-74e8, doi: 10.1126 / science.1175834
- ↑ Giday Wolde-Gabriel et al .: The Geological, Isotopic, Botanical, Invertebrate, and Lower Vertebrate Surroundings of Ardipithecus ramidus. In: Science. Volume 326, 2009, pp. 65, 65e1-65e5, doi: 10.1126 / science.1175817
- ↑ a b Graeme D. Ruxton, David M. Wilkinson: Avoidance of overheating and selection for both hair loss and bipedality in hominins. In: PNAS. Volume 108, No. 52, 2011, pp. 20965-20969, doi: 10.1073 / pnas.1113915108
-
↑ Peter E. Wheeler: The evolution of bipedality and loss of functional body hair in hominids. In: Journal of Human Evolution. Volume 13, No. 1, 1984, pp. 91-98, doi: 10.1016 / S0047-2484 (84) 80079-2
The Naked and the Bipedal. Early hominids may have stood up and gotten naked as a way to cope with heat stress on the sere African savanna. Tim Folger on discovermagazine.com on November 1, 1993 -
↑ Benjamin H. Passey et al .: High-temperature environments of human evolution in East Africa based on bond ordering in paleosol carbonates. In: PNAS. Volume 107, No. 25, 2011, pp. 11245–11249, doi: 10.1073 / pnas.1001824107
East-African human ancestors lived in hot environments, says Caltech-led team. On: eurekalert.org of June 8, 2010. Here it says as a verbatim quote from the co-author of the PNAS study, John Eiler (Robert P. Sharp Professor of Geology and professor of geochemistry at the California Institute of Technology ): “For example , by standing upright, we intercept less direct sunlight than if we were on all fours, and in hot, open environments, the ground and near-surface air can be appreciably hotter than the air a few feet above the ground. So, by standing upright, we are avoiding a high-temperature environment. " - ^ Graeme D. Ruxton and David M. Wilkinson: Thermoregulation and endurance running in extinct hominins: Wheeler's models revisited. In: Journal of Human Evolution. Volume 61, No. 2, 2011, pp. 169-175, doi: 10.1016 / j.jhevol.2011.02.012
- ↑ C. Owen Lovejoy: The Origin of Man. In: Science. Volume 211, 1981, pp. 341-350, doi: 10.1126 / science.211.4480.341
- ↑ to be read in Science , Volume 217, pp. 295-304; critical comments came from Glynn Isaac, Diahan Harley, James W. Wood, Linda D. Wolfe, J. Patrick Gray, John G. Robinson, Leslie S. Lieberman, Elizbeth H. Peters, Rebecca L. Cann and Allan C. Wilson
- ↑ However , this is only documented for Homo erectus : Curtis W. Marean et al .: Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. In: Nature. Volume 449, 2007, pp. 905-908; doi: 10.1038 / nature06204
- ↑ Carsten Niemitz: The secret of the upright walk. Our evolution was different . CH Beck, Munich, 2004, ISBN 3-406-51606-8
- ^ Carsten Niemitz: The evolution of the upright posture and gait - a review and a new synthesis. In: Natural Sciences. Volume 97, No. 3, pp. 241-263 doi: 10.1007 / s00114-009-0637-3 .
- ↑ Algis Kuliukas: Wading for Food: The Driving Force of the Evolution of Bipedalism? In: Nutrition and Health. Volume 16, No. 4, 2002, pp. 267–289, full text ( Memento from December 8, 2015 in the Internet Archive )
- ↑ Ronald J. Clarke , Phillip Tobias : Sterkfontein member 2 foot bones of the oldest South African hominid. In: Science. Volume 269, 1995, p. 521; doi: 10.1126 / science.7624772
- ↑ Katherine K. Whitcome, Liza J. Shapiro, Daniel E. Lieberman: Fetal load and the evolution of lumbar lordosis in bipedal hominins. In: Nature. Volume 450, 2007, pp. 1075-1078, doi: 10.1038 / nature06342
- ↑ 2 figures: [1] and http://www.goatstar.org/lucyhips.jpg ( Memento from December 26, 2010 in the Internet Archive ) Comparison of the pelvis of chimpanzees, Australopithecus and humans
- ^ A b Scott W. Simpson et al .: A Female Homo erectus Pelvis from Gona , Ethiopia. In: Science. Volume 322, No. 5904, 2008, pp. 1089-1092, doi: 10.1126 / science.1163592
- ↑ Narrow pelvis, large heads and the difficult birth in humans. On: idw-online.de from April 21, 2015.
- ↑ Holly M. Dunsworth et al .: Metabolic hypothesis for human altriciality. In: PNAS. Volume 109, No. 38, 2012, pp. 15212–15216, doi: 10.1073 / pnas.1205282109
-
↑ Michael H. Morgan, David R. Carrier: Protective buttressing of the human fist and the evolution of hominin hands. In: Journal of Experimental Biology. Volume 216, 2013, pp. 236-244, doi: 10.1242 / jeb.075713
Fine hands, fists of fury. Our hands evolved for punching, not just dexterity. On: eurekalert.org of December 19, 2012 -
↑ Matthew M. Skinner et al .: Human-like hand use in Australopithecus africanus. In: Science. Volume 347, No. 6220, 2015, pp. 395–399, doi: 10.1126 / science.1261735
Australopithecus africanus - Powerful hands for precise gripping. On: idw-online from January 22, 2015. -
↑ Erin Marie Williams-Hatala et al .: The manual pressures of stone tool behaviors and their implications for the evolution of the human hand. In: Journal of Human Evolution. Volume 119, 2018, pp. 14–26, doi: 10.1016 / j.jhevol.2018.02.008
Eating bone marrow played a key role in the evolution of the human hand. On: eurekalert.org of July 11, 2018 -
↑ Tomoko Sakai et al .: Fetal brain development in chimpanzees versus humans. In: Current Biology. Volume 22, No. 18, 2012, pp. R791-R792, doi: 10.1016 / j.cub.2012.06.062
Human brains outpace chimp brains in the womb. On: eurekalert.org of September 24, 2012
Chimpanzee brains stop growing earlier. ( Memento from October 12, 2012 in the Internet Archive ) On: new-scientist.de from October 8, 2012 (with video) - ↑ Winfried Henke, Hartmut Rothe: Human becoming. Fisherman . Taschenbuch Verlag, Frankfurt am Main 2003, ISBN 3-596-15554-1 , pp. 9 ( fischerverlage.de [PDF] reading sample).
-
↑ Simon Neubauer, Jean-Jacques Hublin and Philipp Gunz: The evolution of modern human brain shape. In: Science Advances . Volume 4, No. 1, 2018, eaao5961, doi: 10.1126 / sciadv.aao5961
Slow and late evolution of the human brain. On: mpg.de from January 24, 2018 - ^ A b Steve Jones et al .: The Cambridge Encyclopedia of Human Evolution. Cambridge University Press, 1992, p. 115
- ↑ a b Axel Kleidon: Climatic constraints on maximum levels of human metabolic activity and their relation to human evolution and global change. In: Climatic Change. Volume 95, No. 3-4, 2009, pp. 405-431, doi: 10.1007 / s10584-008-9537-3
- ↑ Roger S. Seymour et al .: Fossil skulls reveal that blood flow rate to the brain increased faster than brain volume during human evolution. In: Royal Society Open Science. Online publication from August 31, 2016, doi: 10.1098 / rsos.160305
-
^ Leslie C. Aiello, Peter Wheeler: The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution. In: Current Anthropology Volume 36, No. 2, 1995, pp. 199–221, doi: 10.1086 / 204350 , full text (PDF) ( Memento from January 26, 2012 in the Internet Archive ); cf. Expensive Tissue Hypothesis
Ana Navarrete, Carel P. van Schaik and Karin Isler: Energetics and the evolution of human brain size. In: Nature. Volume 480, 2011, pp. 91-93, doi: 10.1038 / nature10629 - ^ David W. Schwartzman and George Middendorf: Biospheric cooling and the emergence of intelligence. In: G. Lemarchand, K. Meech (eds.): A new era in bioastronomy. Volume 213, 2000, ASP Conference Series, pp. 425-429
- ↑ David Schwartzman, George Middendorf, Miranda Armor-Chelu: Was climate the prime releaser for encephalization? An editorial comment. In: Climate Change. Volume 95, No. 3-4, 2009, pp. 439-447, doi: 10.1007 / s10584-009-9603-5
- ^ Drew H. Bailey and David C. Geary: Hominid Brain Evolution. Testing Climatic, Ecological, and Social Competition Models. In: Human Nature. Volume 20, No. 1, 2009, pp. 67-79, doi: 10.1007 / s12110-008-9054-0
- ^ David Robson: A brief history of the brain. In: New Scientist. No. 2831, September 24, 2011, p. 44
-
^ Ronald M. Nowak: Walker's mammals of the world. 6th edition. Johns Hopkins University Press, Baltimore 1999, p. 613
Wolfgang Maier: Primates, Primaten, Herrentiere. In: Wilfried Westheide, Reinhard Rieger (Ed.): Special Zoology. Part 2. Vertebrate or skull animals. Spectrum Academic Publishing House, Heidelberg / Berlin 2004, p. 573 - ↑ brain volume. On: Spektrum.de , accessed on September 10, 2015
- ↑ a b c d e f g h i j k Matthew M. Skinner and Bernard Wood: The evolution of modern human life history - a paleontological perspective. In: Kristen Hawkes and Richard R. Paine (Eds.): The Evolution of Modern Human Life History. School of American Research Press, Santa Fe 2006, p. 351, ISBN 978-1-930618-72-5 . - The arithmetic mean and in brackets the 95% confidence interval are shown .
- ^ Gen Suwa et al .: The Ardipithecus ramidus Skull and Its Implications for Hominid Origins. In: Science. Volume 326, 2009, pp. 68, 68e1-68e7, doi: 10.1126 / science.1175825
- ↑ Gary J. Sawyer, Viktor Deak: The Long Way to Man. Life pictures from 7 million years of evolution. Spektrum Akademischer Verlag, Heidelberg, 2008, p. 79
- ↑ Friedemann Schrenk : The early days of man. The way to Homo sapiens , CH Beck, 1997, p. 70
- ↑ Dean Falk et al .: Brain shape in human microcephalics and Homo floresiensis. In: PNAS. Volume 104, No. 7, 2007, pp. 2513-2518, doi: 10.1073 / pnas.0609185104
- ↑ Hansell H. Stedman et al .: Myosin gene mutation correlates with anatomical changes in the human lineage. In: Nature. Volume 428, 2004, pp. 415-418, doi: 10.1038 / nature02358
- ^ Dean Falk et al .: Metopic suture of Taung (Australopithecus africanus) and its implications for hominin brain evolution. In: PNAS. Volume 109, No. 22, 2012, pp. 8467-8470, doi: 10.1073 / pnas.1119752109 , full text
- ↑ Markus Bastir et al .: Evolution of the base of the brain in highly encephalized human species. In: Nature Communications . Volume 2, Article No. 588, 2011, doi: 10.1038 / ncomms1593
- ^ Emiliano Bruner: The apex of the Incarnation. In: Brain & Mind . No. 10, 2016, p. 44
- ↑ Roger S. Seymour, Vanya Bosiocic and Edward P. Snelling: Fossil skulls reveal that blood flow rate to the brain increased faster than brain volume during human evolution. In: Royal Society Open Science . Online publication from August 31, 2016, doi: 10.1098 / rsos.160305
- ↑ Chet C. Sherwood et al .: Aging of the cerebral cortex differs between humans and chimpanzees. In: PNAS. Volume 108, No. 32, 2011, pp. 13029-13034, doi: 10.1073 / pnas.1016709108
-
↑ Rodrigo S. Lacruz, Chris B. Stringer , William H. Kimbel, Bernard Wood , Katerina Harvati , Paul O'Higgins, Timothy G. Bromage and Juan-Luis Arsuaga : The evolutionary history of the human face. Review Article in: Nature Ecology & Evolution. Volume 3, 2019, pp. 726–736, doi: 10.1038 / s41559-019-0865-7
Need for social skills helped shape modern human face. On: eurekalert.org from April 15, 2019 - ↑ Bernard Wood , Terry Harrison : The evolutionary context of the first hominins. In: Nature. Volume 470, 2011, p. 348, doi: 10.1038 / nature09709
- ↑ Katherine D. Zink and Daniel E. Lieberman: Impact of meat and Lower Palaeolithic food processing techniques on chewing in humans. In: Nature. Volume 531, 2016, pp. 500–503, doi: 10.1038 / nature16990
- ^ Charles Darwin: The Descent of Man, and Selection in Relation to Sex. London, John Murray, 1871, Volume 1, p. 199: “In each great region of the world the living mammals are closely related to the extinct species of the same region. It is therefore probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man's nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere. "
- ↑ Charles Darwin: The Descent of Man and Sexual Selection. 3rd edition, completely revised edition, translated from English by J. Victor Carus. In: Ch. Darwin's collected works, Vol. 5, E. Schweizerbart'sche Verlagshandlung (E. Koch), Stuttgart 1875, p. 68; Digitized
- ^ Gen Suwa et al .: Paleobiological Implications of the Ardipithecus ramidus Dentition. In: Science. Volume 326, 2009, pp. 69, 94-99, doi: 10.1126 / science.1175824
- ^ Gen Suwa et al .: Paleobiological Implications of the Ardipithecus ramidus Dentition. In: Science. Volume 326, No. 5949, 2009, p. 69, doi: 10.1126 / science.1175824
- ↑ a b Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, pp. 33–34
- ↑ Friedemann Schrenk in: Spectrum of Science . No. 9/2010, p. 69. - According to the same source, for a Homo erectus from Indonesia and for Homo rudolfensis , it has been proven that their front teeth still had two tooth roots.
- ↑ Christopher Dean: Developmental sequences and rates of growth in tooth length in hominoids. In: RJ Radlanski and H. Renz (Eds.): Structure, Function and Evolution of Teeth. Proceedings of the 10th International Symposium on Dental Morphology. M Marketing Services Brünne GbR, Berlin 1995, pp. 308-313
- ^ Gabriele A. Macho and Bernard Wood: The role of time and timing in hominid dental evolution. In: Evolutionary Anthropology. Volume 4, No. 1, 1995, pp. 17–31, doi: 10.1002 / evan.1360040105 , full text (PDF; 2.5 MB) ( Memento from February 2, 2014 in the Internet Archive )
- ^ Alan Mann, Michelle Lampl and Janet Monge: Patterns of ontogeny in human evolution: Evidence from dental development. In: American Journal of Physical Anthropology. Volume 33, Supplement S11 (= Yearbook of Physical Anthropology), 1990, pp. 111-150, doi: 10.1002 / ajpa.1330330507
- ^ Glenn C. Conroy and Michael W. Vannier: Dental development in South African australopithecines. Part I: Problems of pattern and chronology. In: American Journal of Physical Anthropology. Volume 86, No. 2, 1991, pp. 121-136, doi: 10.1002 / ajpa.1330860204
- ↑ Noreen von Cramon-Taubadel: Global human mandibular variation reflects differences in agricultural and hunter-gatherer subsistence strategies. In: PNAS. Volume 108, No. 49, 2011, pp. 19546–19551, doi: 10.1073 / pnas.1113050108
- ↑ HV Weems and TR Fasulo: Human Lice: Body Lice, Pediculus humanus humanus Linnaeus and Head Louse, Pediculus humanus capitis De Geer (Insecta: Phthiraptera (= Anoplura): Pediculidae). Document EENY-104 of the US Department of Entomology and Nematology, as of March 2017, full text (PDF; 510 kB) .
- ^ DL Reed, JE Light, JM Allen, JJ Kirchman: Pair of lice lost or parasites regained: the evolutionary history of anthropoid primate lice. In: BMC biology. Volume 5, No. 7, 2007, doi: 10.1186 / 1741-7007-5-7 , PMID 17343749 , PMC 1828715 (free full text).
- ↑ a b Ralf Kittler, Manfred Kayser and Mark Stoneking: Molecular Evolution of Pediculus humanus and the Origin of Clothing. In: Current Biology. Volume 13, No. 16, 2003, pp. 1414-1417, doi: 10.1016 / S0960-9822 (03) 00507-4
- ↑ a b Melissa A. Toups et al .: Origin of Clothing Lice Indicates Early Clothing Use by Anatomically Modern Humans in Africa. In: Molecular Biology and Evolution. Volume 28, No. 1, 2011, pp. 29-32, doi: 10.1093 / molbev / msq234
- ↑ Alan. R. Rogers et al .: Genetic Variation at the MC1R Locus and the Time since Loss of Human Body Hair. In: Current Anthropology. Volume 45, No. 1, 2004, pp. 105-108, doi: 10.1086 / 381006
- ↑ Thomas Junker: The evolution of man. CH Beck, Munich 2008, p. 89
- ↑ a b Nina G. Jablonski : Why people are naked. In: Spektrum der Wissenschaft , 10/2010, pp. 60–67
- ↑ Mark Pagel and Walter Bodmer: A naked ape would have fewer parasites. In: Proceedings of the Royal Society (London) B. Volume 270, No. Suppl 1, 2003, pages S117-S119, doi: 10.1098 / rsbl.2003.0041
- ^ Yana G. Kamberov et al .: Modeling Recent Human Evolution in Mice by Expression of a Selected EDAR Variant. In: Cell . Volume 152, No. 4, 2013, pp. 691-702, doi: 10.1016 / j.cell.2013.01.016
- ↑ Friedemann Schrenk , Stephanie Müller: The 101 most important questions: primeval times. CH Beck, Munich 2006, p. 49
- ↑ Inge Kronberg: Geographical distribution of skin color. In: Biology in Our Time. March 29, 2006, pp. 76-77
- ↑ Ephemeral complexion. In: Geo. No. 10, 2009, p. 22
- ↑ literally: "Skin coloration in humans is adaptive and labile. Skin pigmentation levels have changed more than once in human evolution. Because of this, skin coloration is of no value in determining phylogenetic relationships among modern human groups. "Quoted from: Nina G. Jablonski, George Chaplin: The evolution of human skin coloration. In: Journal of Human Evolution. Volume 39, 2000, pp. 57–106 (here: p. 57), doi: 10.1006 / jhev.2000.0403 , full text (PDF; 3.2 MB) ( Memento from September 6, 2015 in the Internet Archive )
- ↑ Prof. Dieter Böning: Sexual selection in humans - a taboo topic? In: Spectrum of Science. Issue 12/2010, p. 7
- ^ Richard Leakey, The origin of humankind , p. 20
- ^ Cade McCall and Tania Singer : The animal and human neuroendocrinology of social cognition, motivation and behavior. In: Nature Neuroscience. Volume 15, 2012, pp. 681-688, doi: 10.1038 / nn.3084
-
↑ Mary Ann Raghanti et al .: A neurochemical hypothesis for the origin of hominids. In: PNAS. Volume 115, No. 6, E1108-E1116O, 2018, doi: 10.1073 / pnas.1719666115
Dopamine may have given humans our social edge over other apes. On: sciencemag.org of January 22, 2018 - ↑ Jeremy DeSilva, Julie Lesnik: Chimpanzee neonatal brain size: Implications for brain growth in Homo erectus. In: Journal of Human Evolution. Volume 51, No. 2, 2006, pp. 207-212, doi: 10.1016 / j.jhevol.2006.05.006
- ↑ a b Hélène Coqueugniot et al .: Early brain growth in Homo erectus and implications for cognitive ability. In: Nature. Volume 431, 2004, pp. 299–302, doi: 10.1038 / nature02852 , full text (PDF)
- ^ Ann Gibbons: The Birth of Childhood. In: Science. Volume 322, No. 5904, 2008, pp. 1040-1043, doi: 10.1126 / science.322.5904.1040 ; Image from Science : Comparison of the pelvic shape of Australopithecus , Homo erectus and humans
- ^ Tanya M. Smith et al .: Earliest evidence of modern human life history in North African early Homo sapiens. In: PNAS. Volume 104, No. 15, 2007, pp. 6128-6133, doi: 10.1073 / pnas.0700747104
- ↑ Timothy G. Bromage and M. Christopher Dean: Re-evaluation of the age at death of immature fossil hominids. In: Nature. Volume 317, 1985, pp. 525-527, doi: 10.1038 / 317525a0
- ^ Tanya M. Smith et al .: Dental evidence for ontogenetic differences between modern humans and Neanderthals. In: PNAS. Volume 107, No. 49, 2010, pp. 20923-20928, doi: 10.1073 / pnas.1010906107
- ↑ Average life expectancy in industrialized countries
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, p. 39
-
↑ Caleb E. Finch: Evolution of the human lifespan and diseases of aging: Roles of infection, inflammation, and nutrition. In: PNAS. Volume 107, Supplement 1, 2010, pp. 1718-1724, doi: 10.1073 / pnas.0909606106
Why we outlive our ape ancestors. On: eurekalert.org of December 2, 2009 - ↑ Craig Packer et al .: Reproductive cessation in female mammals. In: Nature. Volume 392, 1998, pp. 807-811, doi: 10.1038 / 33910
- ↑ Alison Motlok: Bye-bye babies. In: New Scientist, December 13, 2008, pp. 41-43
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, p. 191
- ↑ Mirkka Lahdenperä et al .: Fitness benefits of prolonged post-reproductive lifespan in women. In: Nature. Volume 428, 2004, pp. 178-181, doi: 10.1038 / nature02367
- ↑ a b Erwin J. Haeberle: The sexuality of man. Manual and atlas. 2nd, expanded edition, Verlag Walter de Gruyter, Berlin and New York 1985, p. 140, ISBN 3-11-010694-9
- ^ Larry J. Young: Being Human: Love: Neuroscience reveals all. In: Nature. Volume 457, 2009, p. 148, doi: 10.1038 / 457148a
- ↑ a b c d C. Owen Lovejoy : The Origin of Man. In: Science . Volume 211, 1981, pp. 341-350, doi: 10.1126 / science.211.4480.341
- ^ AF Dixson: Baculum length and copulatory behavior in primates. In: American Journal of Primatology. Volume 13, No. 1, 1987, pp. 51-60, doi: 10.1002 / ajp.1350130107
- ↑ Damian Labuda et al .: Female-to-Male Breeding Ratio in Modern Humans - an Analysis Based on Historical Recombinations. In: The American Journal of Human Genetics. Volume 86, No. 3, 2010, pp. 353-363, doi: 10.1016 / j.ajhg.2010.01.029
-
^ Asif A. Ghazanfar et al .: Cineradiography of Monkey Lip-Smacking Reveals Putative Precursors of Speech Dynamics. In: Current Biology. Volume 22, No. 13, 2012, pp. 1176–1182, doi: 10.1016 / j.cub.2012.04.055
Current Biology: Monkey lip smacking offers insights into the evolution of language. On: idw-online from May 31, 2012 - ↑ a b c Winfried Henke, Hartmut Rothe: Tribal history of humans. Springer Verlag, Berlin 1999, pp. 80-81
- ^ A b W. Tecumseh Fitch: The evolution of language. In: New Scientist. December 4, 2010, Supplement Instant Expert 6 , pp. I – viii
-
↑ Speech gene: Why people can talk. On: aerzteblatt.de of November 12, 2009:
Human language genes are unique. On: berliner-zeitung.de from August 15, 2002 -
↑ Wolfgang Enard et al .: Molecular evolution of FOXP2, a gene involved in speech and language. In: Nature. Volume 418, 2002, pp. 869-872, doi: 10.1038 / nature01025
A mutation separates humans and monkeys. On: spiegel.de from August 15, 2002 - ↑ Roger Fouts, Stephen Tukel Mills: Our Nearest Relatives. Learn from chimpanzees what it means to be human. Limes Verlag, Munich 1998, ISBN 3-8090-3013-9 (also published as a paperback by Droemer Knaur in 2002, ISBN 3-426-77420-8 )
- ↑ Grammar out of nowhere. About deaf and dumb children and creole languages. P. 257 In: Dieter E. Zimmer: Experiments of life. Zurich: Haffmans Verlag. 1989 ISBN 3-251-00139-6
- ↑ Steven Pinker : The Language Instinct. Munich: Kindler 1996, ISBN 3-463-40267-X
- ↑ Time to speak chimp. About language experiments with great apes. In: Dieter E. Zimmer: Experiments of Life. Zurich: Haffmans Verlag. 1989 ISBN 3-251-00139-6
- ↑ "Increasing evidence indicates that vocal signals can have a similar function to grooming in mediating social interactions and relationships." Nathalie C. Greeno, Stuart Semple: Sex differences in vocal communication among adult rhesus macaques. In: Evolution and Human Behavior. Volume 30, No. 2, 2009, pp. 141-145, doi: 10.1016 / j.evolhumbehav.2008.09.002
- ↑ Eörs Szathmáry, Szabolcs Számadó: Language: a social history of words. In: Nature. Volume 456, 2008, pp. 40-41, doi: 10.1038 / 456040a
-
^ Samuel Bowles: Conflict: Altruism's midwife. In: Nature. Volume 456, 2008, pp. 326–327, doi: 10.1038 / 456326a , full text (PDF; 144 kB)
Martin A. Nowak: Generosity: A winner's advice. In: Nature. Volume 456, 2008, p. 579, doi: 10.1038 / 456579a - ↑ Otto König : Culture and behavior research - introduction to cultural ethology . DTV-Verlag 1970. ISBN 3-423-00614-5
- ↑ Richard Dawkins: Meme, The New Replicators. In: The Selfish Gene (original: The Selfish Gene , Oxford University Press, 1976). Anniversary edition 2007, pp. 316–334. ISBN 3-499-19609-3 .
- ↑ Susan Blackmore : The Power of Memes. Heidelberg, Berlin: Spektrum Akademischer Verlag, 2000, ISBN 3-8274-1601-9 .
- ↑ Kate Douglas: Culture club. In: New Scientist. November 20, 2010, pp. 38-41
- ↑ Andrew Whiten, Jane Goodall et al .: Cultures in chimpanzees. In: Nature. Volume 399, 1999, pp. 682-685, doi: 10.1038 / 21415
- ^ Luke Rendell, Hal Whitehead: Culture in whales and dolphins. In: Behavioral and Brain Sciences. Volume 24, 2001, pp. 309-324, doi: 10.1017 / S0140525X0100396X
- ↑ Carel P. van Schaik et al .: Orangutan Cultures and the Evolution of Material Culture. In: Science. Volume 299, No. 5603, 2003, pp. 102-105, doi: 10.1126 / science.1078004
- ↑ Susan Perry et al .: Social Conventions in Wild White ‐ faced Capuchin Monkeys: Evidence for Traditions in a Neotropical Primate. In: Current Anthropology. Volume 44, No. 2, 2003, pp. 241-268
- ↑ Doron M. Behar et al .: The Dawn of Human Matrilineal Diversity. In: American Journal of Human Genetics. Volume 82, 2008, pp. 1130-1140, doi: 10.1016 / j.ajhg.2008.04.002
- ↑ Quentin D. Atkinson: Phonemic Diversity Supports a Serial Founder Effect Model of Language Expansion from Africa. In: Science. Volume 332, No. 6027, 2011, pp. 346-349, doi: 10.1126 / science.1199295
- ↑ Caleb Everett et al .: Climate, vocal folds, and tonal languages: Connecting the physiological and geographic dots. In: PNAS. Volume 112, No. 5, 2015, pp. 1322-1327, doi: 10.1073 / pnas.1417413112
- ↑ Murray Gell-Mann , Merritt Ruhlen : The origin and evolution of word order. In: PNAS. Volume 108, No. 42, 2011, pp. 17290-17295, doi: 10.1073 / pnas.1113716108
- ^ 'I the dog see', our ancestors said. In: New Scientist, October 15, 2011, p. 19
-
↑ newscientist.com of February 17, 2010: “The writing on the cave wall.” Also published under the title Messages from the stone age in New Scientist , No. 2748 of February 20, 2010, pp. 32-34
diepresse.com dated February 19, 2010: “Archeology: The writing on the wall. The cave paintings do not only consist of representations, but perhaps also of signs. " - ↑ Sherwood L. Washburn : Tools and human evolution. In: Scientific American. Volume 203, 1960, pp. 63-75
- ↑ Jane Goodall: Tool-Using and Aimed Throwing in a Community of Free-Living Chimpanzees. In: Nature. Volume 201, 1964, pp. 1264-1266, doi: 10.1038 / 2011264a0
- ^ William C. McGrew: Chimpanzee Technology. In: Science. Volume 328, No. 5978, 2010, pp. 579-580, doi: 10.1126 / science.1187921
- ↑ Michael Haslam et al .: Primate archeology. In: Nature. Volume 460, 2009, pp. 339-344, doi: 10.1038 / nature08188
- ↑ S. Semaw, P. Renne, JWK Harris, CS Feibel, RL Bernor, N. Fesseha and K. Mowbray: 2.5-million-year-old stone tools from Gona, Ethiopia. In: Nature. Volume 385, 1997, pp. 333-336 doi: 10.1038 / 385333a0
- ↑ a b Sileshi Semaw: The World's Oldest Stone Artefacts from Gona, Ethiopia: Their Implications for Understanding Stone Technology and Patterns of Human Evolution Between 2.6-1.5 Million Years Ago. In: Journal of Archaeological Science. Volume 27, 2000, pp. 1197-1214 doi: 10.1006 / jasc.1999.0592
- ↑ Shannon P. McPherron et al .: Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika. In: Nature. Volume 466, 2010, pp. 857-860, doi: 10.1038 / nature09248
- ↑ Manuel Domínguez-Rodrigo et al .: Configurational approach to identifying the earliest hominin butchers. In: PNAS. Volume 107, No. 49, 2010, pp. 20929-20934, doi: 10.1073 / pnas.1013711107
-
↑ Jayne Wilkins et al .: Evidence for Early Hafted Hunting Technology. In: Science . Volume 338, No. 6109, 2012, pp. 942–946, doi: 10.1126 / science.1227608
eurekalert.org (with ill.) From November 15, 2012: Archaeologists identify spear tips used in hunting a half-million years ago. - ↑ a b David R. Braun et al .: Early hominin diet included diverse terrestrial and aquatic animals 1.95 Ma in East Turkana, Kenya. In: PNAS. Volume 107, No. 22, 2010, pp. 10002-10007, doi: 10.1073 / pnas.1002181107
-
↑ Anders Högberg and Lars Larsson: Lithic technology and behavioral modernity: New results from the Still Bay site, Hollow Rock Shelter , Western Cape Province, South Africa. In: Journal of Human Evolution. Volume 61, No. 2, 2011, pp. 133–155, doi: 10.1016 / j.jhevol.2011.02.006
eurektalert.com of June 21, 2011: Cutting edge training developed the human brain 80,000 years ago. - ^ Aldo Faisal et al .: The Manipulative Complexity of Lower Paleolithic Stone Toolmaking. In: PLoS ONE. Volume 5, No. 11: e13718, 2010, doi: 10.1371 / journal.pone.0013718
-
^ Kyle S. Brown et al .: Fire As an Engineering Tool of Early Modern Humans. In: Science. Volume 325, No. 5942, 2009, pp. 859–862, doi: 10.1126 / science.1175028
eurekalert.org of August 13, 2009: Early modern humans use fire to engineer tools from stone. -
↑ Pat Shipman: The Animal Connection and Human Evolution. In: Current Anthropology. Volume 51, No. 4, 2010, pp. 519-538, short version
eurekalert.org of July 20, 2010: New hypothesis for human evolution and human nature. -
↑ Eliso Kvavadze et al .: 30,000-Year-Old Wild Flax Fibers. In: Science. Volume 325, 2009, p. 1359, doi: 10.1126 / science.1175404
eurekalert.org of September 10, 2009: Archaeologists discover oldest-known fiber materials used by early humans. - ↑ James M. Adovasio et al .: Perishable Industries from Dolni Vestonice I: New Insights into the Nature and Origin of the Gravettian. In: Archeology, Ethnology and Anthropology of Eurasia. Volume 2, No. 6, pp. 48-65
- ^ Hansjürgen Müller-Beck: The Ice Ages. Natural history and human history. CH Beck, Munich 2005, p. 96, ISBN 3-406-50863-4
-
↑ Ken Sayers, C. Owen Lovejoy : Blood, Bulbs, and Bunodonts: On Evolutionary Ecology and the Diets of Ardipithecus, Australopithecus, and Early Homo. In: The Quarterly Review of Biology. Volume 89, No. 4, 2014, pp. 319-357, doi: 10.1086 / 678568
What was the 'Paleo diet'? There was far more than one, study suggests. On: eurekalert.org of December 16, 2014. - ^ Peter S. Ungar, Matt Sponheimer: The Diets of Early Hominins. In: Science. Volume 334, No. 6053, 2011, pp. 190–193, doi: 10.1126 / science.1207701
- ↑ Chris Organ et al .: Phylogenetic rate shifts in feeding time during the evolution of Homo. In: PNAS. Volume 108, No. 35, 2011, pp. 14555-14559, doi: 10.1073 / pnas.1107806108
- ↑ Gary J. Sawyer, Viktor Deak: The Long Way to Man. Life pictures from 7 million years of evolution . Spectrum , Heidelberg 2008, ISBN 978-3-8274-1915-6 , p. 36 .
- ^ Ann Gibbons: Food for Thought. In: Science. Volume 316. No. 5831, 2007, pp. 1558-1560, doi: 10.1126 / science.316.5831.1558
- ^ Felix Warneken and Alexandra G. Rosati: Cognitive capacities for cooking in chimpanzees. In: Proceedings of the Royal Society B. Volume 282, No. 1809, 2015, doi: 10.1098 / rspb.2015.0229
- ↑ Naama Goren-Inbar, Nira Alperson, Mordechai E. Kislev, Orit Simchoni, Yoel Melamed, Adi Ben-Nun and Ella Werker: Evidence of Hominin Control of Fire at Gesher Benot Ya`aqov, Israel. In: Science. Volume 304, 2004, pp. 725-727, doi: 10.1126 / science.1095443
- ^ Wil Roebroeks , Paola Villa: On the earliest evidence for habitual use of fire in Europe. In: PNAS. Volume 108, No. 13, 2011, pp. 5209-5214, doi: 10.1073 / pnas.1018116108
- ↑ The connection between food processing by heating and the evolution of Homo sapiens is discussed in detail in a book by Richard Wrangham: Catching Fire. How cooking made us human - a new theory of human evolution. DVA, Munich 2009, ISBN 978-3-421-04399-3 (Original title: Catching Fire: How Cooking Made Us Human. )
- ^ Neil Mann, Meat in the human diet: An anthropological perspective . In: Nutrition & Dietetics . tape 64 , s4, 2007, p. S102 – S107 , doi : 10.1111 / j.1747-0080.2007.00194.x .
- ^ Ann Gibbons: What's for Dinner? Researchers Seek Our Ancestors' Answers. In: Science. Volume 326, No. 5959, 2009, pp. 1478-1479, doi: 10.1126 / science.326.5959.1478
- ↑ Frank Eckhardt: Back to the roots! The original human diet . In: Utz Thimm, Karl-Heinz Wellmann (Ed.): On everyone's lips. Diet today . Suhrkamp, Frankfurt am Main 2004, ISBN 3-518-45602-4 , pp. 27-37 .
- ^ Ann Gibbons: How the modern body shaped up. In: Science. Volume 336, No. 6081, 2012, pp. 538-539, doi: 10.1126 / science.336.6081.538-b
- ↑ Christopher S. Henshilwood et al .: Emergence of Modern Human Behavior: Middle Stone Age Engravings from South Africa. In: Science. Volume 295, 2002, pp. 1278-1280, doi: 10.1126 / science.1067575 . See also the illustration in The Japan Times of January 13, 2002
- ↑ Abdeljalil Bouzouggar et al .: 82,000-year-old shell beads from North Africa and implications for the origins of modern human behavior. In: PNAS . Volume 104, No. 24, 2007, pp. 9964-9969 doi: 10.1073 / pnas.0703877104
- ↑ Pierre-Jean Texier et al .: A Howiesons Poort tradition of engraving ostrich eggshell containers dated to 60,000 years ago at Diepkloof Rock Shelter, South Africa. In: PNAS. Volume 107, No. 14, 2010, pp. 6180-6185, doi: 10.1073 / pnas.0913047107
- ↑ David Lordkipanidze , Abesalom Vekua et al .: The earliest toothless hominin skull. In: Nature. Volume 434, 2005, pp. 717-718, doi: 10.1038 / 434717b ISSN 0028-0836
- ↑ Alejandro Bonmatí et al .: Middle Pleistocene lower back and pelvis from an aged human individual from the Sima de los Huesos site, Spain. In: PNAS. Volume 107, No. 43, 2010, pp. 18386-18391, doi: 10.1073 / pnas.1012131107
- ↑ Charles Darwin: The Descent of Man and Sexual Selection. Volume 1, p. 120. In the original wording: "There is no evidence that man was aboriginally endowed with the ennobling belief in the existence of an Omnipotent God." Charles Darwin: The Descent of Man, and Selection in Relation to Sex. Volume 1, p. 65
- ↑ Charles Darwin: The Descent of Man and Sexual Selection. Volume 1, p. 121. In the original wording: “As soon as the important faculties of the imagination, wonder, and curiosity, together with some power of reasoning, had become partially developed, man would naturally have craved to understand what was passing around him , and have vaguely speculated on his own existence. "Charles Darwin: The Descent of Man, and Selection in Relation to Sex. Volume 1, p. 65
- ↑ Ara Norenzayan, Azim F. Shariff: The Origin and Evolution of Religious Prosociality. In: Science. Volume 322, No. 5898, 2008, pp. 58-62, doi: 10.1126 / science.1158757
- ↑ Ilkka Pyysiäinen and Marc Hauser: The origins of religion: evolved adaptation or by-product? In: Trends in Cognitive Sciences. Volume 14, No. 3, 2010, pp. 104-109, doi: 10.1016 / j.tics.2009.12.007
- ↑ Pascal Boyer: Being human: Religion: Bound to believe? In: Nature. Volume 455, 2008, pp. 1038-1039, doi: 10.1038 / 4551038a
- ↑ Michael Brooks: Natural born believers. In: New Scientist. Volume 201, No. 2694, 2009, pp. 30-33, doi: 10.1016 / S0262-4079 (09) 60372-9
- ↑ Elizabeth Culotta: On the Origin of Religion. In: Science. Volume 326, No. 5954, 2009, pp. 784-787, doi: 10.1126 / science. 326 784
- ↑ Michael Balter: On the Origin of Art and Symbolism. In: Science. Volume 323, No. 5915, 2009, pp. 709-711, doi: 10.1126 / science.323.5915.709
- ↑ Eduardo Palacio-Pérez: Cave art and the theory of art: the origins of the religious interpretation of Palaeolithic Graphics Expression. In: Oxford Journal of Archeology. Volume 29, No. 1, 2010, pp. 1-14, doi: 10.1111 / j.1468-0092.2009.00337.x . A summary can be found at eurekalert.org of March 26, 2010: Religious beliefs are the basis of the origins of Palaeolithic art.
- ↑ a b Vilayanur S. Ramachandran , Sandra Blakeslee: The blind woman who can see. Puzzling phenomena of our consciousness . Rowohlt, Reinbek bei Hamburg 2002, p. 285
![[1]](http://anthro.palomar.edu/hominid/images/pelvis_and_feet.gif){kind=link}
{kind=link}
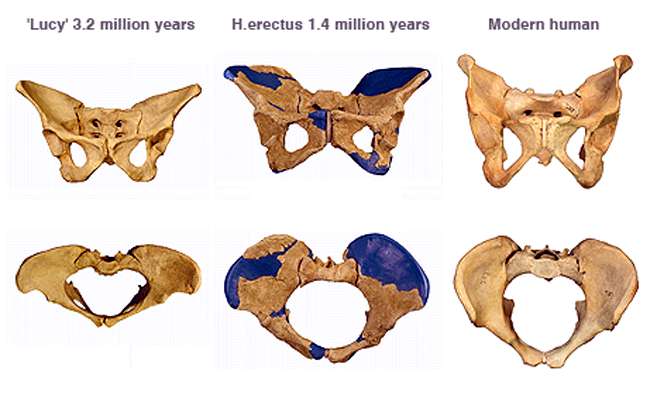{kind=link}